

Dynamic plasticity of large-scale chromatin structure revealed by self-assembly of engineered chromosome regions
- PMID: 20819934
- PMCID: PMC2935575
- DOI: 10.1083/jcb.200912167
Dynamic plasticity of large-scale chromatin structure revealed by self-assembly of engineered chromosome regions
Abstract
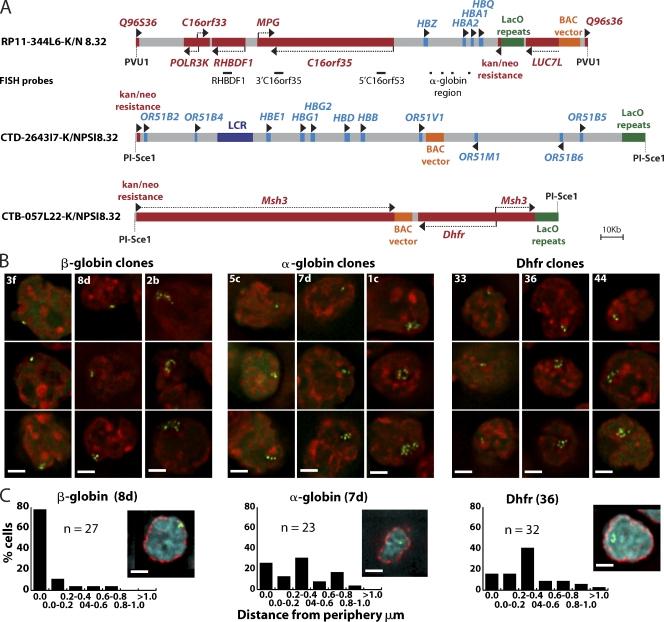
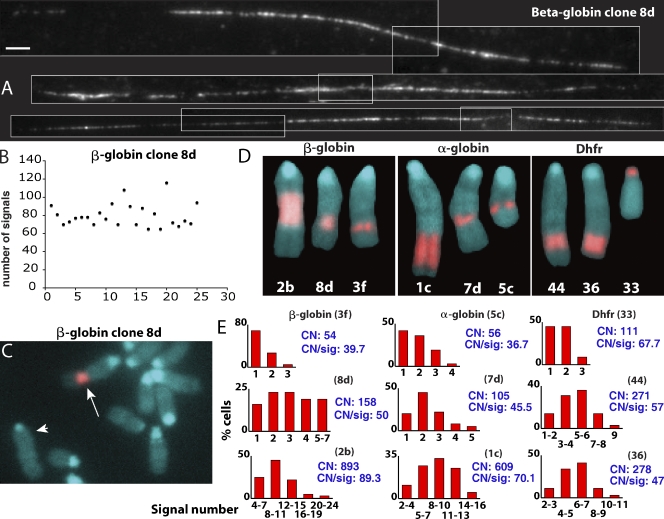
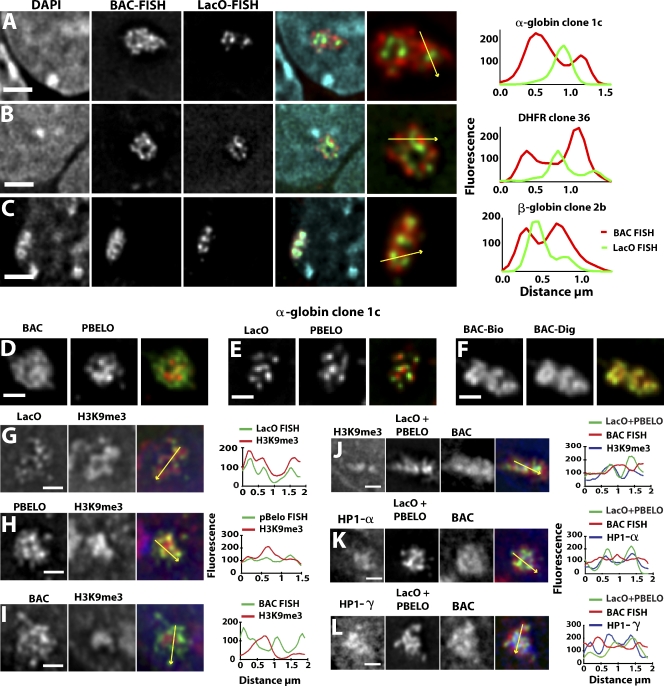
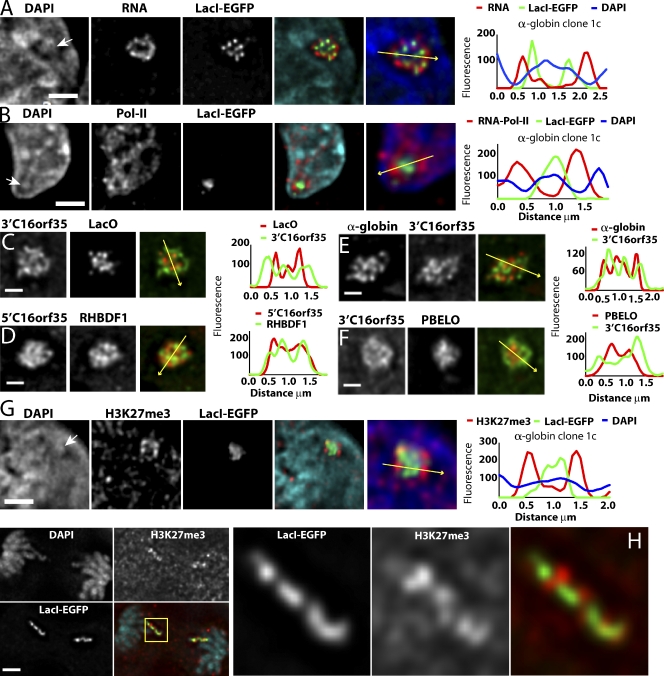
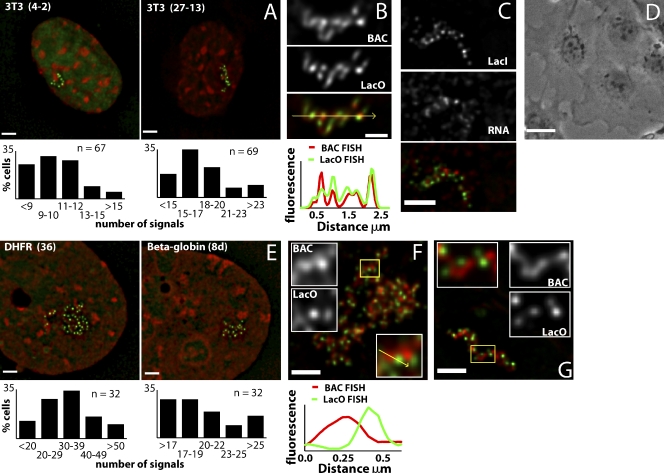
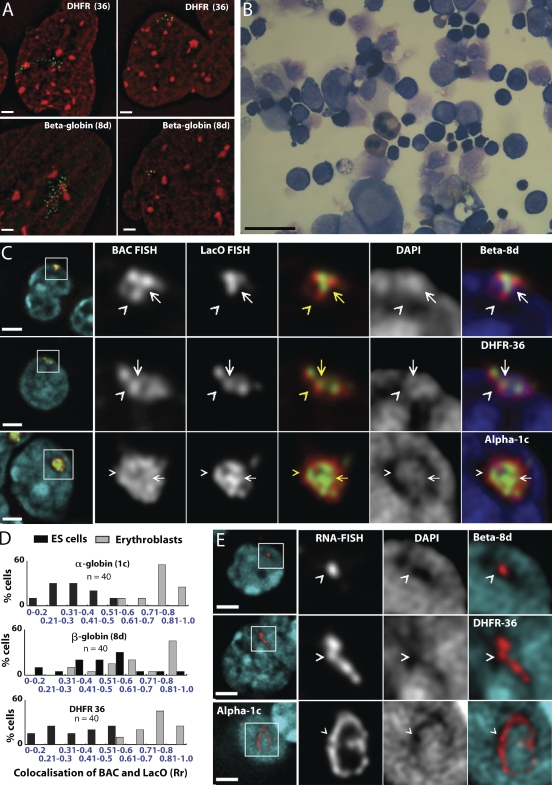
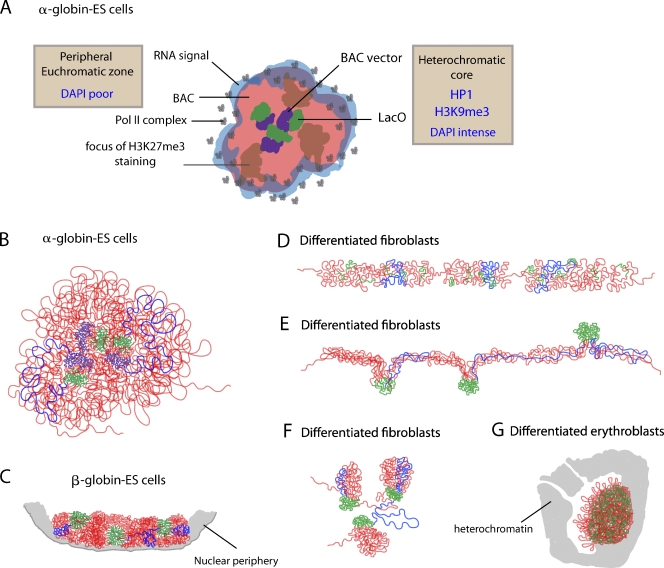
Interphase chromatin compaction well above the 30-nm fiber is well documented, but the structural motifs underlying this level of chromatin folding remain unknown. Taking a reductionist approach, we analyzed in mouse embryonic stem (ES) cells and ES-derived fibroblasts and erythroblasts the folding of 10-160-megabase pair engineered chromosome regions consisting of tandem repeats of bacterial artificial chromosomes (BACs) containing approximately 200 kilobases of mammalian genomic DNA tagged with lac operator (LacO) arrays. Unexpectedly, linear mitotic and interphase chromatid regions formed from noncontiguously folded DNA topologies. Particularly, in ES cells, these model chromosome regions self-organized with distant sequences segregating into functionally distinct, compact domains. Transcriptionally active and histone H3K27me3-modified regions positioned toward the engineered chromosome subterritory exterior, with LacO repeats and the BAC vector backbone localizing within an H3K9me3, HP1-enriched core. Differential compaction of Dhfr and alpha- and beta-globin transgenes was superimposed on dramatic, lineage-specific reorganization of large-scale chromatin folding, demonstrating a surprising plasticity of large-scale chromatin organization.
Figures
References
-
- Beatty B.G., Scherer S.W. 2002. Human chromosome mapping of single copy genes. FISH: A Practical Approach. Beatty B., Mai S., Squire J., Oxford University Press, Oxford: 29–53
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
