

Functional dichotomy of ribosomal proteins during the synthesis of mammalian 40S ribosomal subunits
- PMID: 20819938
- PMCID: PMC2935573
- DOI: 10.1083/jcb.201005117
Functional dichotomy of ribosomal proteins during the synthesis of mammalian 40S ribosomal subunits
Abstract
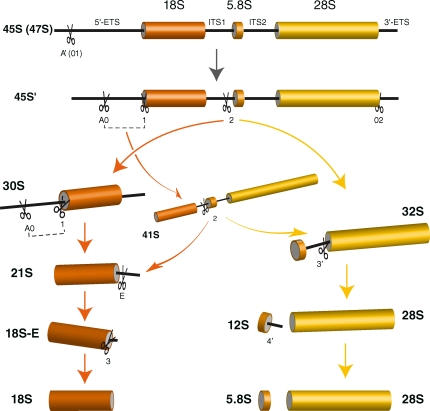
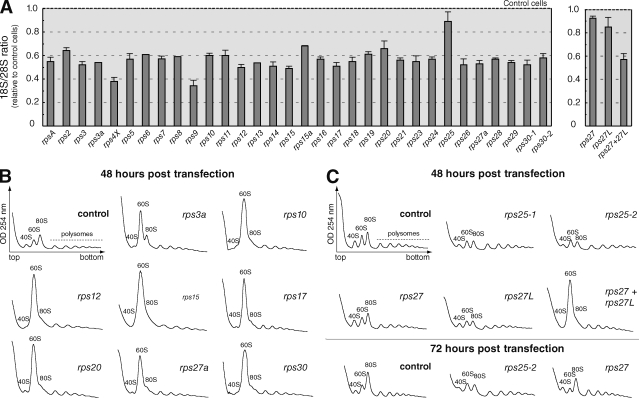
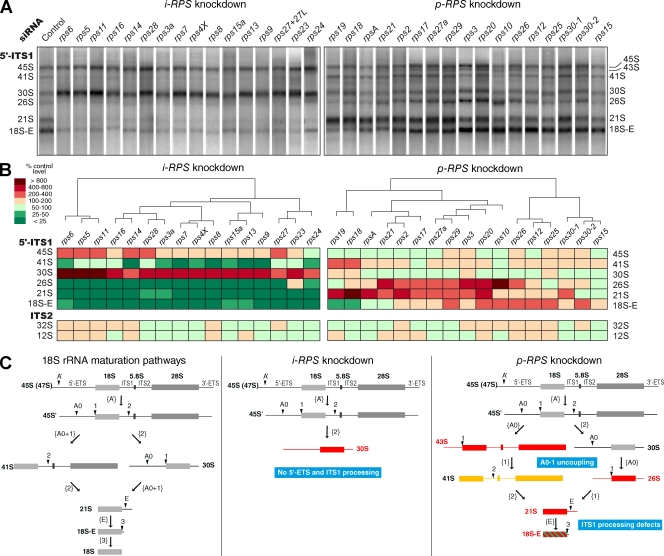
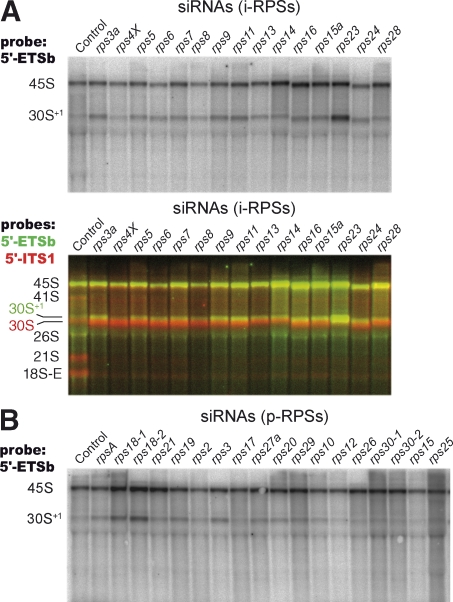
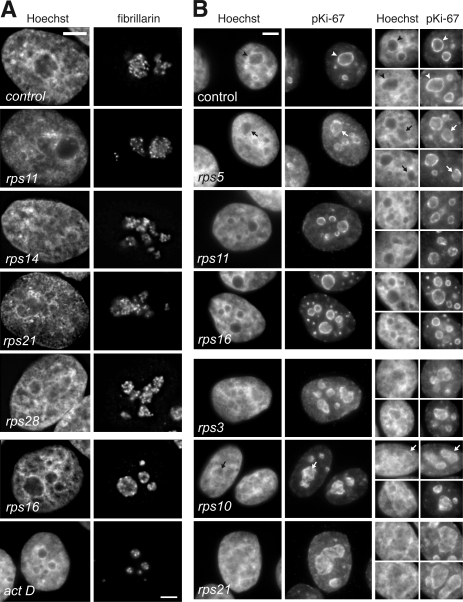
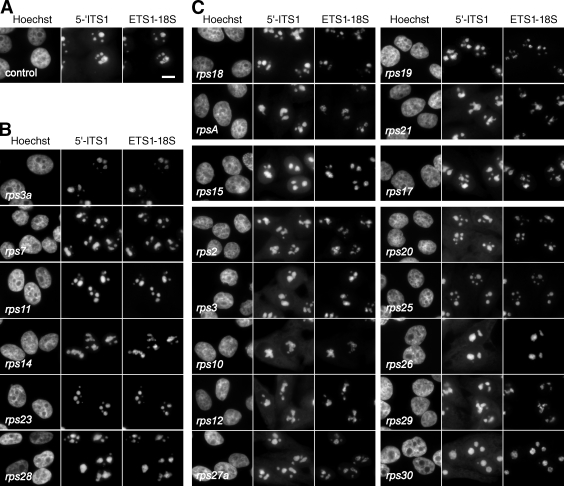
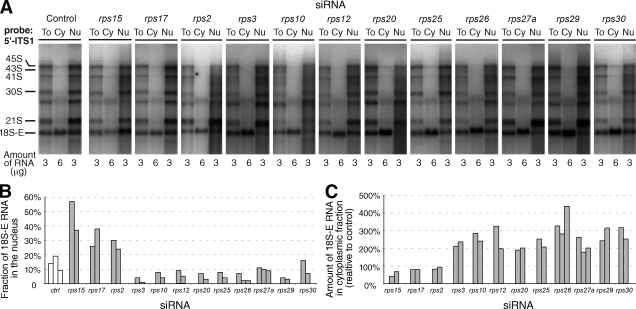
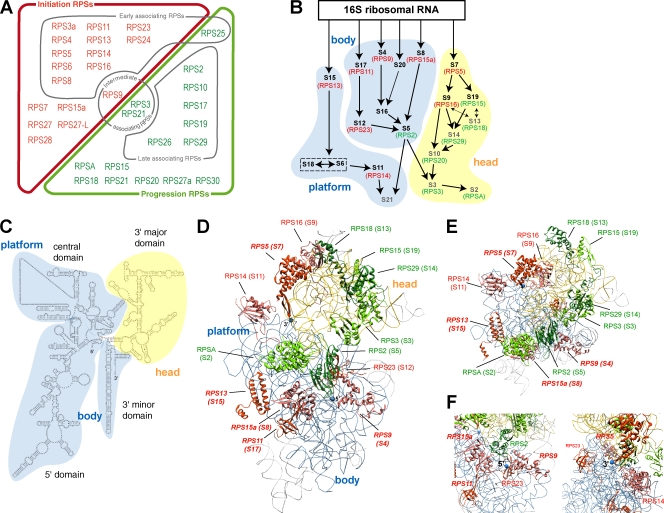
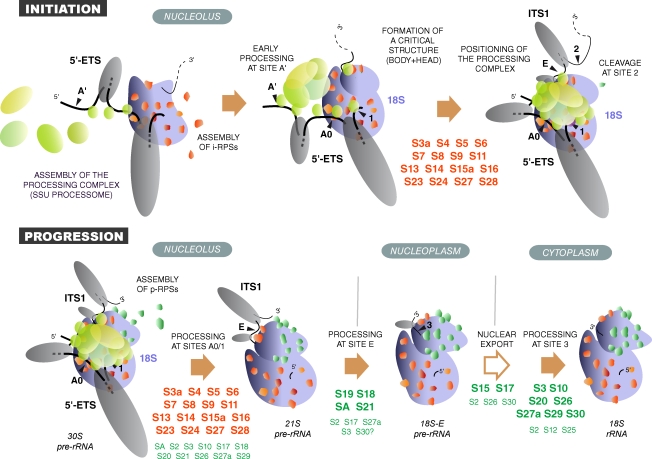
Our knowledge of the functions of metazoan ribosomal proteins in ribosome synthesis remains fragmentary. Using siRNAs, we show that knockdown of 31 of the 32 ribosomal proteins of the human 40S subunit (ribosomal protein of the small subunit [RPS]) strongly affects pre-ribosomal RNA (rRNA) processing, which often correlates with nucleolar chromatin disorganization. 16 RPSs are strictly required for initiating processing of the sequences flanking the 18S rRNA in the pre-rRNA except at the metazoan-specific early cleavage site. The remaining 16 proteins are necessary for progression of the nuclear and cytoplasmic maturation steps and for nuclear export. Distribution of these two subsets of RPSs in the 40S subunit structure argues for a tight dependence of pre-rRNA processing initiation on the folding of both the body and the head of the forming subunit. Interestingly, the functional dichotomy of RPS proteins reported in this study is correlated with the mutation frequency of RPS genes in Diamond-Blackfan anemia.
Figures
Similar articles
-
Impaired ribosome biogenesis in Diamond-Blackfan anemia.Blood. 2007 Feb 1;109(3):1275-83. doi: 10.1182/blood-2006-07-038372. Epub 2006 Oct 19. Blood. 2007. PMID: 17053056 Free PMC article.
-
Cells depleted for RPS19, a protein associated with Diamond Blackfan Anemia, show defects in 18S ribosomal RNA synthesis and small ribosomal subunit production.Blood Cells Mol Dis. 2007 Jul-Aug;39(1):35-43. doi: 10.1016/j.bcmd.2007.02.001. Epub 2007 Mar 21. Blood Cells Mol Dis. 2007. PMID: 17376718
-
Nuclear export competence of pre-40S subunits in fission yeast requires the ribosomal protein Rps2.Nucleic Acids Res. 2008 Nov;36(19):6132-42. doi: 10.1093/nar/gkn625. Epub 2008 Sep 27. Nucleic Acids Res. 2008. PMID: 18820293 Free PMC article.
-
Nuclear export and cytoplasmic maturation of ribosomal subunits.FEBS Lett. 2007 Jun 19;581(15):2783-93. doi: 10.1016/j.febslet.2007.05.013. Epub 2007 May 11. FEBS Lett. 2007. PMID: 17509569 Review.
-
Pre-ribosomes on the road from the nucleolus to the cytoplasm.Trends Cell Biol. 2003 May;13(5):255-63. doi: 10.1016/s0962-8924(03)00054-0. Trends Cell Biol. 2003. PMID: 12742169 Review.
Cited by
-
Structure of a human pre-40S particle points to a role for RACK1 in the final steps of 18S rRNA processing.Nucleic Acids Res. 2016 Sep 30;44(17):8465-78. doi: 10.1093/nar/gkw714. Epub 2016 Aug 16. Nucleic Acids Res. 2016. PMID: 27530427 Free PMC article.
-
Involvement of human ribosomal proteins in nucleolar structure and p53-dependent nucleolar stress.Nat Commun. 2016 Jun 6;7:11390. doi: 10.1038/ncomms11390. Nat Commun. 2016. PMID: 27265389 Free PMC article.
-
Targeting the Extra-Cellular Matrix-Tumor Cell Crosstalk for Anti-Cancer Therapy: Emerging Alternatives to Integrin Inhibitors.Front Oncol. 2020 Jul 22;10:1231. doi: 10.3389/fonc.2020.01231. eCollection 2020. Front Oncol. 2020. PMID: 32793493 Free PMC article. Review.
-
RIOK2 phosphorylation by RSK promotes synthesis of the human small ribosomal subunit.PLoS Genet. 2021 Jun 14;17(6):e1009583. doi: 10.1371/journal.pgen.1009583. eCollection 2021 Jun. PLoS Genet. 2021. PMID: 34125833 Free PMC article.
-
Biochemical and clinical effects of RPS20 expression in renal clear cell carcinoma.Oncol Rep. 2023 Jan;49(1):22. doi: 10.3892/or.2022.8459. Epub 2022 Dec 9. Oncol Rep. 2023. PMID: 36484407 Free PMC article.
References
-
- Auger-Buendia M.A., Longuet M., Tavitian A. 1979. Kinetic studies on ribosomal proteins assembly in preribosomal particles and ribosomal subunits of mammalian cells. Biochim. Biophys. Acta. 563:113–128 - PubMed
-
- Barkić M., Crnomarkovic S., Grabusić K., Bogetić I., Panić L., Tamarut S., Cokarić M., Jerić I., Vidak S., Volarević S. 2009. The p53 tumor suppressor causes congenital malformations in Rpl24-deficient mice and promotes their survival. Mol. Cell. Biol. 29:2489–2504 10.1128/MCB.01588-08 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
