

Cotransport of water by the Na+-K+-2Cl(-) cotransporter NKCC1 in mammalian epithelial cells
- PMID: 20819947
- PMCID: PMC3002443
- DOI: 10.1113/jphysiol.2010.194738
Cotransport of water by the Na+-K+-2Cl(-) cotransporter NKCC1 in mammalian epithelial cells
Abstract
Water transport by the Na+-K+-2Cl(-) cotransporter (NKCC1) was studied in confluent cultures of pigmented epithelial (PE) cells from the ciliary body of the fetal human eye. Interdependence among water, Na+ and Cl(-) fluxes mediated by NKCC1 was inferred from changes in cell water volume, monitored by intracellular self-quenching of the fluorescent dye calcein. Isosmotic removal of external Cl(-) or Na+ caused a rapid efflux of water from the cells, which was inhibited by bumetanide (10 μm). When returned to the control solution there was a rapid water influx that required the simultaneous presence of external Na+ and Cl(-). The water influx could proceed uphill, against a transmembrane osmotic gradient, suggesting that energy contained in the ion fluxes can be transferred to the water flux. The influx of water induced by changes in external [Cl(-)] saturated in a sigmoidal fashion with a Km of 60 mm, while that induced by changes in external [Na+] followed first order kinetics with a Km of about 40 mm. These parameters are consistent with ion transport mediated by NKCC1. Our findings support a previous investigation, in which we showed water transport by NKCC1 to be a result of a balance between ionic and osmotic gradients. The coupling between salt and water transport in NKCC1 represents a novel aspect of cellular water homeostasis where cells can change their volume independently of the direction of an osmotic gradient across the membrane. This has relevance for both epithelial and symmetrical cells.
Figures
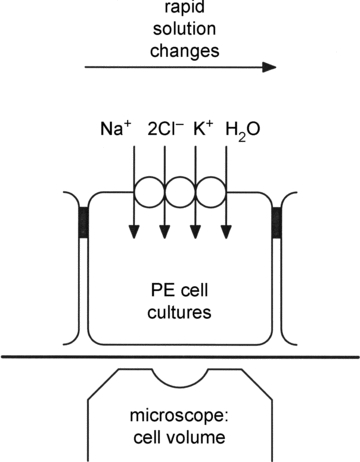




 ) for each pulse was estimated from the initial rates of cell swelling using eqn (2). Each data point represents the mean ±
) for each pulse was estimated from the initial rates of cell swelling using eqn (2). Each data point represents the mean ± ) for each pulse was estimated from the initial rates of cell swelling using eqn (2). Each data point represents the mean ±
) for each pulse was estimated from the initial rates of cell swelling using eqn (2). Each data point represents the mean ±




Comment in
-
Water cotransport in pigmented epithelial cells.J Physiol. 2010 Nov 1;588(Pt 21):4063-4. doi: 10.1113/jphysiol.2010.199521. J Physiol. 2010. PMID: 21037312 Free PMC article. No abstract available.
-
NKCC1 cotransporters: keeping an 'ion' them.J Physiol. 2011 Feb 15;589(Pt 4):781-2. doi: 10.1113/jphysiol.2010.203919. J Physiol. 2011. PMID: 21486847 Free PMC article. Review. No abstract available.
References
-
- Agre P, Nielsen S, Ottersen OP. Towards a molecular understanding of water homeostasis in the brain. Neuroscience. 2004;12:849–850. - PubMed
-
- Alvarez-Leefmans FJ. Chloride transporters in presynaptic inhibition, pain and neurogenic inflammation. In: Alvarez-Leefmans FJ, Delpire E, editors. Physiology and Pathology of Chloride Transporters and Channels in the Nervous System: From Molecules to Diseases. San Diego, CA: Elsevier-Academic Press; 2009.
-
- Alvarez-Leefmans FJ, Altamirano J, Crowe WE. Use of ion-selective microelectrodes and fluorescent probes to measure cell volume. Methods Neurosci. 1995;27:361–391.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources