

Histone deacetylase 2 is required for chromatin condensation and subsequent enucleation of cultured mouse fetal erythroblasts
- PMID: 20823130
- PMCID: PMC2995558
- DOI: 10.3324/haematol.2010.029827
Histone deacetylase 2 is required for chromatin condensation and subsequent enucleation of cultured mouse fetal erythroblasts
Abstract
Background: During the final stages of differentiation of mammalian erythroid cells, the chromatin is condensed and enucleated. We previously reported that Rac GTPases and their downstream target, mammalian homolog of Drosophila diaphanous 2 (mDia2), are required for enucleation of in vitro cultured mouse fetal liver erythroblasts. However, it is not clear how chromatin condensation is achieved and whether it is required for enucleation.
Design and methods: Mouse fetal liver erythroblasts were purified from embryonic day 14.5 pregnant mice and cultured in erythropoietin-containing medium. Enucleation was determined by flow-cytometry based analysis after treatment with histone deacetylase inhibitors or infection with lentiviral short hairpin RNA.
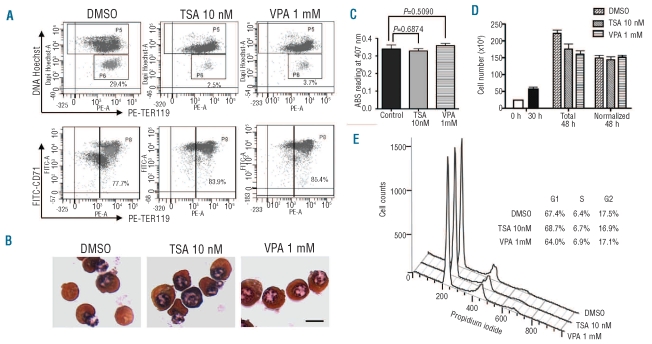
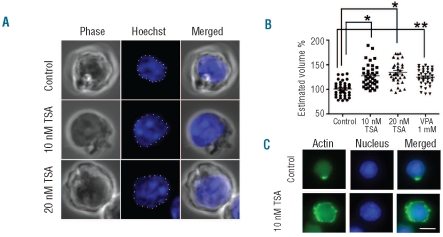
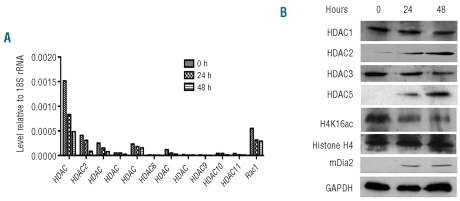
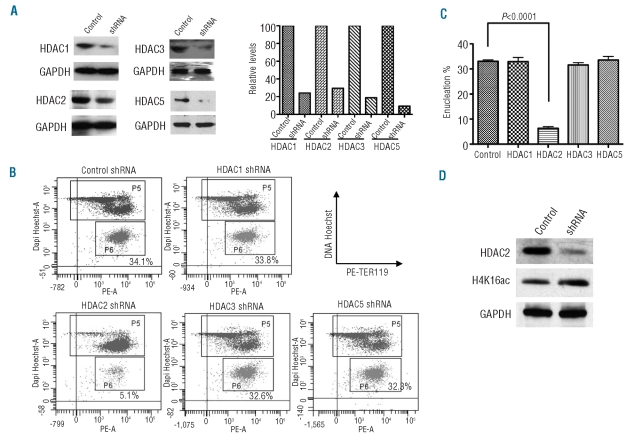
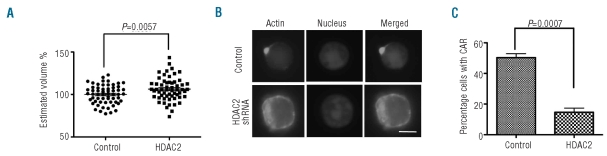
Results: We showed that histone deacetylases play critical roles in chromatin condensation and enucleation in cultured mouse fetal liver erythroblasts. Enzymatic inhibition of histone deacetylases by trichostatin A or valproic acid prior to the start of enucleation blocked chromatin condensation, contractile actin ring formation and enucleation. We further demonstrated that histone deacetylases 1, 2, 3 and 5 are highly expressed in mouse fetal erythroblasts. Short hairpin RNA down-regulation of histone deacetylase 2, but not of the other histone deacetylases, phenotypically mimicked the effect of trichostatin A or valproic acid treatment, causing significant inhibition of chromatin condensation and enucleation. Importantly, knock-down of histone deacetylase 2 did not affect erythroblast proliferation, differentiation, or apoptosis.
Conclusions: These results identify histone deacetylase 2 as an important regulator, mediating chromatin condensation and enucleation in the final stages of mammalian erythropoiesis.
Figures

Comment in
-
Erythroblast enucleation.Haematologica. 2010 Dec;95(12):1985-8. doi: 10.3324/haematol.2010.033225. Haematologica. 2010. PMID: 21123437 Free PMC article. No abstract available.
References
-
- Richmond TD, Chohan M, Barber DL. Turning cells red: signal transduction mediated by erythropoietin. Trends Cell Biol. 2005;15(3):146–55. - PubMed
-
- Ji P, Jayapal SR, Lodish HF. Enucleation of cultured mouse fetal erythroblasts requires Rac GTPases and mDia2. Nat Cell Biol. 2008;10(3):314–21. - PubMed
-
- Yoshida H, Kawane K, Koike M, Mori Y, Uchiyama Y, Nagata S. Phosphatidylserine-dependent engulfment by macrophages of nuclei from erythroid precursor cells. Nature. 2005;437(7059):754–8. - PubMed
-
- Chasis JA, Prenant M, Leung A, Mohandas N. Membrane assembly and remodeling during reticulocyte maturation. Blood. 1989;74(3):1112–20. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
