

Viral terminal protein directs early organization of phage DNA replication at the bacterial nucleoid
- PMID: 20823229
- PMCID: PMC2944753
- DOI: 10.1073/pnas.1010530107
Viral terminal protein directs early organization of phage DNA replication at the bacterial nucleoid
Abstract
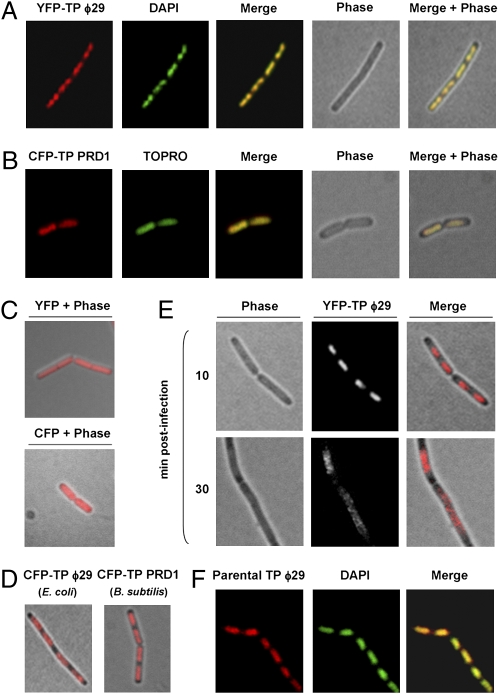
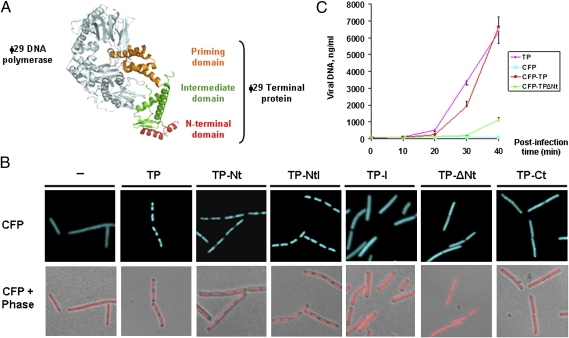
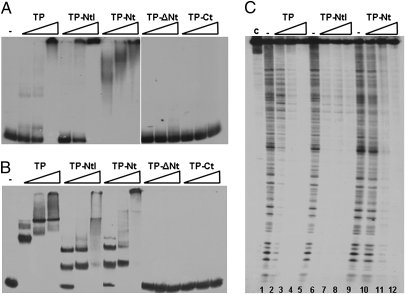
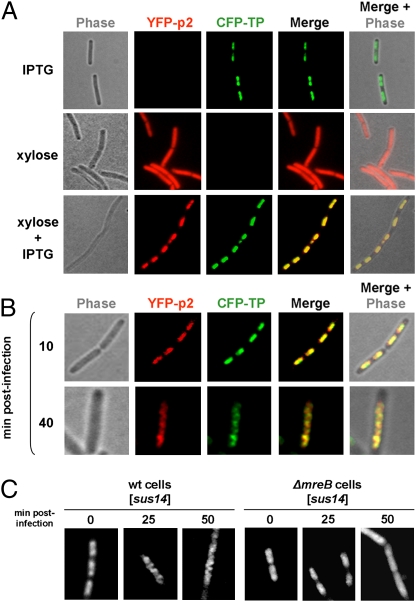
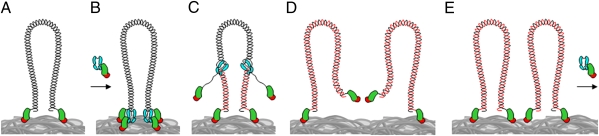
The mechanism leading to protein-primed DNA replication has been studied extensively in vitro. However, little is known about the in vivo organization of the proteins involved in this fundamental process. Here we show that the terminal proteins (TPs) of phages ϕ29 and PRD1, infecting the distantly related bacteria Bacillus subtilis and Escherichia coli, respectively, associate with the host bacterial nucleoid independently of other viral-encoded proteins. Analyses of phage ϕ29 revealed that the TP N-terminal domain (residues 1-73) possesses sequence-independent DNA-binding capacity and is responsible for its nucleoid association. Importantly, we show that in the absence of the TP N-terminal domain the efficiency of ϕ29 DNA replication is severely affected. Moreover, the TP recruits the phage DNA polymerase to the bacterial nucleoid, and both proteins later are redistributed to enlarged helix-like structures in an MreB cytoskeleton-dependent way. These data disclose a key function for the TP in vivo: organizing the early viral DNA replication machinery at the cell nucleoid.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
The actin-like MreB cytoskeleton organizes viral DNA replication in bacteria.Proc Natl Acad Sci U S A. 2009 Aug 11;106(32):13347-52. doi: 10.1073/pnas.0906465106. Proc Natl Acad Sci U S A. 2009. PMID: 19654094 Free PMC article.
-
New insights in the ϕ29 terminal protein DNA-binding and host nucleoid localization functions.Mol Microbiol. 2014 Jan;91(2):232-41. doi: 10.1111/mmi.12456. Epub 2013 Nov 27. Mol Microbiol. 2014. PMID: 24205926
-
Dissecting the role of the ϕ29 terminal protein DNA binding residues in viral DNA replication.Nucleic Acids Res. 2015 Mar 11;43(5):2790-801. doi: 10.1093/nar/gkv127. Epub 2015 Feb 26. Nucleic Acids Res. 2015. PMID: 25722367 Free PMC article.
-
Protein-Primed Replication of Bacteriophage Φ29 DNA.Enzymes. 2016;39:137-67. doi: 10.1016/bs.enz.2016.03.005. Epub 2016 May 12. Enzymes. 2016. PMID: 27241929 Review.
-
Role of host factors in bacteriophage φ29 DNA replication.Adv Virus Res. 2012;82:351-83. doi: 10.1016/B978-0-12-394621-8.00020-0. Adv Virus Res. 2012. PMID: 22420858 Review.
Cited by
-
DNA-Binding Proteins Essential for Protein-Primed Bacteriophage Φ29 DNA Replication.Front Mol Biosci. 2016 Aug 5;3:37. doi: 10.3389/fmolb.2016.00037. eCollection 2016. Front Mol Biosci. 2016. PMID: 27547754 Free PMC article. Review.
-
Dual role of φ29 DNA polymerase Lys529 in stabilisation of the DNA priming-terminus and the terminal protein-priming residue at the polymerisation site.PLoS One. 2013 Sep 4;8(9):e72765. doi: 10.1371/journal.pone.0072765. eCollection 2013. PLoS One. 2013. PMID: 24023769 Free PMC article.
-
My life with bacteriophage phi29.J Biol Chem. 2012 Dec 28;287(53):44568-79. doi: 10.1074/jbc.X112.433458. Epub 2012 Nov 2. J Biol Chem. 2012. PMID: 23124207 Free PMC article.
-
Bacteriophage protein Gp46 is a cross-species inhibitor of nucleoid-associated HU proteins.Proc Natl Acad Sci U S A. 2022 Mar 1;119(9):e2116278119. doi: 10.1073/pnas.2116278119. Proc Natl Acad Sci U S A. 2022. PMID: 35193978 Free PMC article.
-
Phage 29 phi protein p1 promotes replication by associating with the FtsZ ring of the divisome in Bacillus subtilis.Proc Natl Acad Sci U S A. 2013 Jul 23;110(30):12313-8. doi: 10.1073/pnas.1311524110. Epub 2013 Jul 8. Proc Natl Acad Sci U S A. 2013. PMID: 23836667 Free PMC article.
References
-
- Salas M. Protein-priming of DNA replication. Annu Rev Biochem. 1991;60:39–71. - PubMed
-
- Salas M. Mechanisms of initiation of linear DNA replication in prokaryotes. Genet Eng (N Y) 1999;21:159–171. - PubMed
-
- de Jong RN, van der Vliet PC, Brenkman AB. Adenovirus DNA replication: Protein priming, jumping back and the role of the DNA binding protein DBP. Curr Top Microbiol Immunol. 2003;272:187–211. - PubMed
-
- Chaconas G, Chen CW. In: The Bacterial Chromosome. Higgins NP, editor. Washington, DC: American Society for Microbiology Press; 2005. pp. 525–539.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
