

The somatic reproductive tissues of C. elegans promote longevity through steroid hormone signaling
- PMID: 20824162
- PMCID: PMC2930862
- DOI: 10.1371/journal.pbio.1000468
The somatic reproductive tissues of C. elegans promote longevity through steroid hormone signaling
Abstract
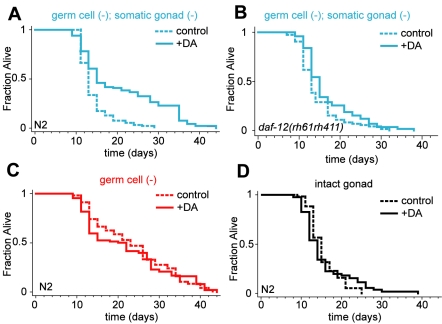
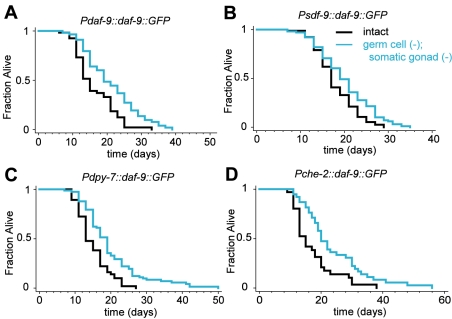
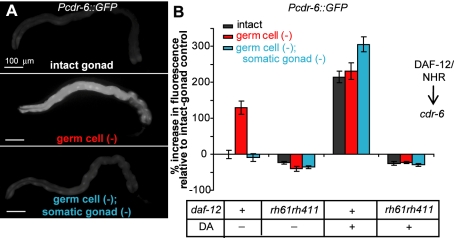
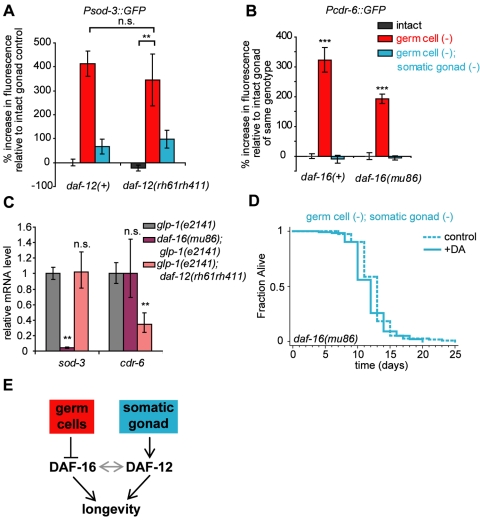
In Caenorhabditis elegans and Drosophila melanogaster, removing the germline precursor cells increases lifespan. In worms, and possibly also in flies, this lifespan extension requires the presence of somatic reproductive tissues. How the somatic gonad signals other tissues to increase lifespan is not known. The lifespan increase triggered by loss of the germ cells is known to require sterol hormone signaling, as reducing the activity of the nuclear hormone receptor DAF-12, or genes required for synthesis of the DAF-12 ligand dafachronic acid, prevents germline loss from extending lifespan. In addition to sterol signaling, the FOXO transcription factor DAF-16 is required to extend lifespan in animals that lack germ cells. DAF-12/NHR is known to assist with the nuclear accumulation of DAF-16/FOXO in these animals, yet we find that loss of DAF-12/NHR has little or no effect on the expression of at least some DAF-16/FOXO target genes. In this study, we show that the DAF-12-sterol signaling pathway has a second function to activate a distinct set of genes and extend lifespan in response to the somatic reproductive tissues. When germline-deficient animals lacking somatic reproductive tissues are given dafachronic acid, their expression of DAF-12/NHR-dependent target genes is restored and their lifespan is increased. Together, our findings indicate that in C. elegans lacking germ cells, the somatic reproductive tissues promote longevity via steroid hormone signaling to DAF-12.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Fatty acid desaturation links germ cell loss to longevity through NHR-80/HNF4 in C. elegans.PLoS Biol. 2011 Mar;9(3):e1000599. doi: 10.1371/journal.pbio.1000599. Epub 2011 Mar 15. PLoS Biol. 2011. PMID: 21423649 Free PMC article.
-
Distinct activities of the germline and somatic reproductive tissues in the regulation of Caenorhabditis elegans' longevity.Genetics. 2008 Jan;178(1):513-26. doi: 10.1534/genetics.107.083253. Genetics. 2008. PMID: 18202391 Free PMC article.
-
A steroid hormone that extends the lifespan of Caenorhabditis elegans.Aging Cell. 2007 Feb;6(1):87-94. doi: 10.1111/j.1474-9726.2006.00268.x. Aging Cell. 2007. PMID: 17266678
-
A pathway that links reproductive status to lifespan in Caenorhabditis elegans.Ann N Y Acad Sci. 2010 Aug;1204:156-62. doi: 10.1111/j.1749-6632.2010.05640.x. Ann N Y Acad Sci. 2010. PMID: 20738286 Review.
-
Steroid hormone regulation of C. elegans and Drosophila aging and life history.Exp Gerontol. 2011 Feb-Mar;46(2-3):141-7. doi: 10.1016/j.exger.2010.08.021. Epub 2010 Sep 17. Exp Gerontol. 2011. PMID: 20854888 Review.
Cited by
-
Roles for ROS and hydrogen sulfide in the longevity response to germline loss in Caenorhabditis elegans.Proc Natl Acad Sci U S A. 2016 May 17;113(20):E2832-41. doi: 10.1073/pnas.1524727113. Epub 2016 May 2. Proc Natl Acad Sci U S A. 2016. PMID: 27140632 Free PMC article.
-
No Significant Increase in the Δ4- and Δ7-Dafachronic Acid Concentration in the Long-Lived glp-1 Mutant, nor in the Mutants Defective in Dauer Formation.G3 (Bethesda). 2015 May 12;5(7):1473-9. doi: 10.1534/g3.115.018812. G3 (Bethesda). 2015. PMID: 25971936 Free PMC article.
-
Regulation of Age-related Decline by Transcription Factors and Their Crosstalk with the Epigenome.Curr Genomics. 2018 Sep;19(6):464-482. doi: 10.2174/1389202919666180503125850. Curr Genomics. 2018. PMID: 30258277 Free PMC article. Review.
-
Characterization of the endogenous DAF-12 ligand and its use as an anthelmintic agent in Strongyloides stercoralis.Elife. 2021 Dec 7;10:e73535. doi: 10.7554/eLife.73535. Elife. 2021. PMID: 34874004 Free PMC article.
-
Fatty acid desaturation links germ cell loss to longevity through NHR-80/HNF4 in C. elegans.PLoS Biol. 2011 Mar;9(3):e1000599. doi: 10.1371/journal.pbio.1000599. Epub 2011 Mar 15. PLoS Biol. 2011. PMID: 21423649 Free PMC article.
References
-
- Williams G. C. Natural selection, the costs of reproduction, and a refinement of lack's principle. The American Naturalist. 1966;100:687.
-
- Hsin H, Kenyon C. Signals from the reproductive system regulate the lifespan of C. elegans. Nature. 1999;399:362–366. - PubMed
-
- Arantes-Oliveira N, Apfeld J, Dillin A, Kenyon C. Regulation of life-span by germ-line stem cells in Caenorhabditis elegans. Science. 2002;295:502–505. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
