

A role for PDGF signaling in expansion of the extra-embryonic endoderm lineage of the mouse blastocyst
- PMID: 20826533
- PMCID: PMC2947752
- DOI: 10.1242/dev.050864
A role for PDGF signaling in expansion of the extra-embryonic endoderm lineage of the mouse blastocyst
Abstract
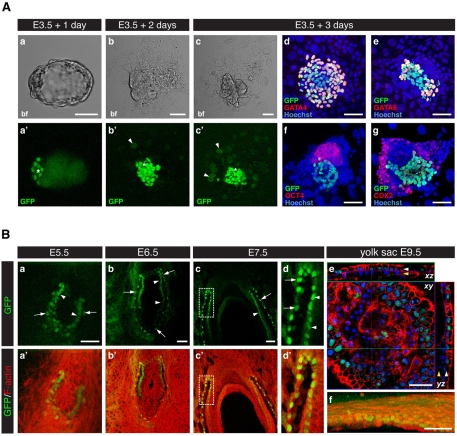
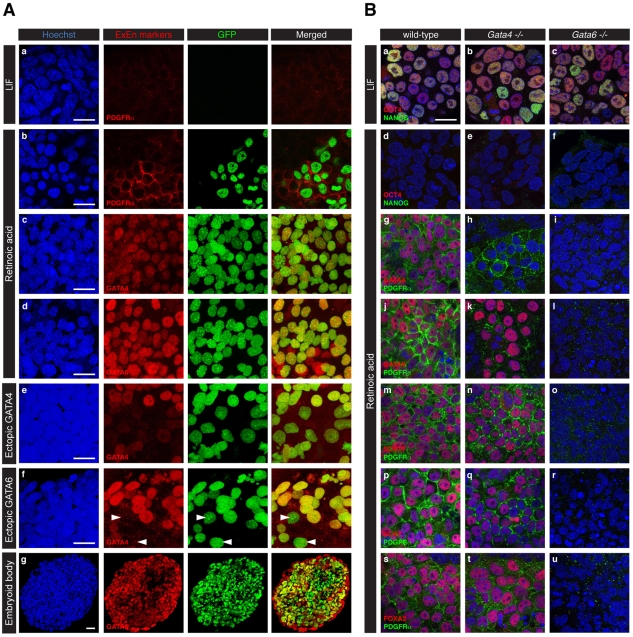
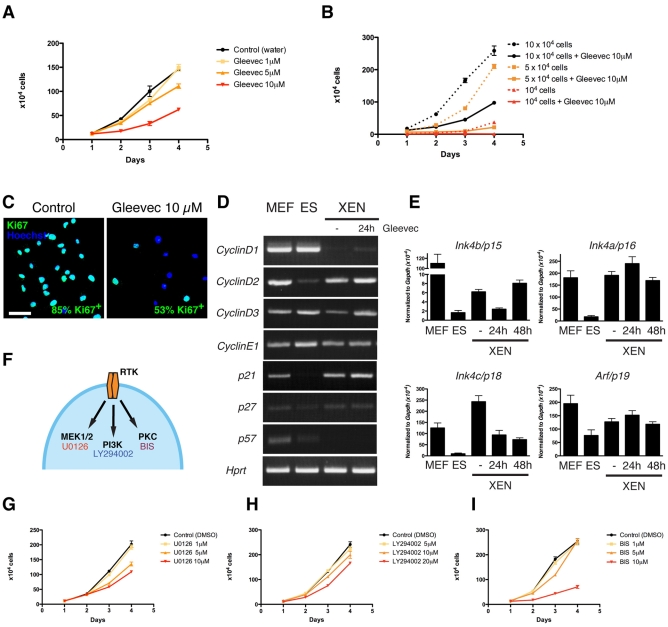
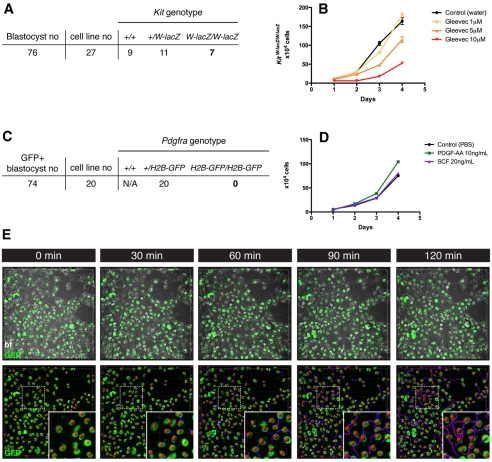
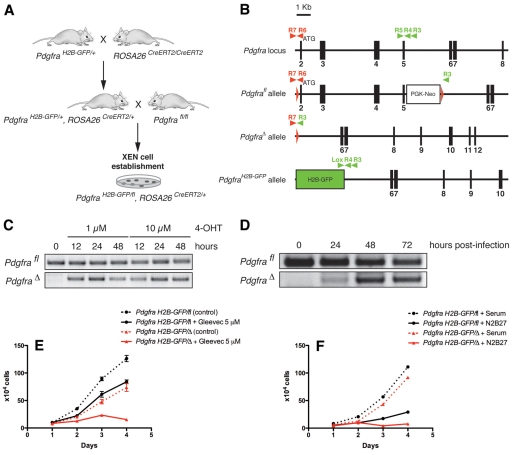
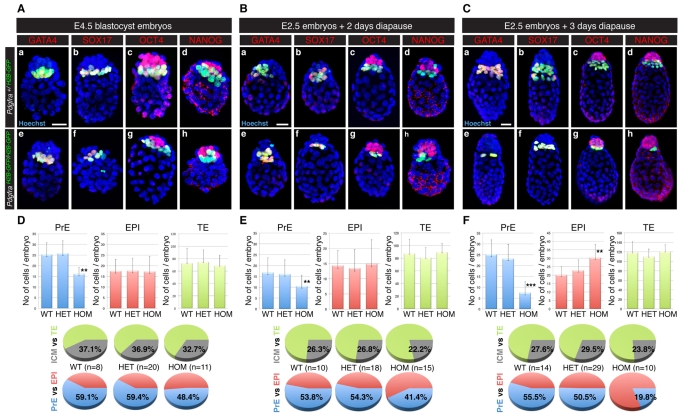
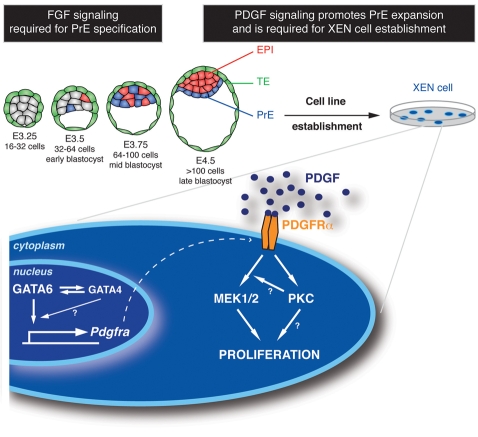
The inner cell mass (ICM) of the implanting mammalian blastocyst comprises two lineages: the pluripotent epiblast (EPI) and primitive endoderm (PrE). We have identified platelet-derived growth factor receptor alpha (PDGFRα) as an early marker of the PrE lineage and its derivatives in both mouse embryos and ex vivo paradigms of extra-embryonic endoderm (ExEn). By combining live imaging of embryos and embryo-derived stem cells expressing a histone H2B-GFP fusion reporter under the control of Pdgfra regulatory elements with the analysis of lineage-specific markers, we found that Pdgfra expression coincides with that of GATA6, the earliest expressed transcriptional regulator of the PrE lineage. We show that GATA6 is required for the activation of Pdgfra expression. Using pharmacological inhibition and genetic inactivation we addressed the role of the PDGF pathway in the PrE lineage. Our results demonstrate that PDGF signaling is essential for the establishment, and plays a role in the proliferation, of XEN cells, which are isolated from mouse blastocyst stage embryos and represent the PrE lineage. Implanting Pdgfra mutant blastocysts exhibited a reduced number of PrE cells, an effect that was exacerbated by delaying implantation. Surprisingly, we also noted an increase in the number of EPI cells in implantation-delayed Pdgfra-null mutants. Taken together, our data suggest a role for PDGF signaling in the expansion of the ExEn lineage. Our observations also uncover a possible role for the PrE in regulating the size of the pluripotent EPI compartment.
Figures
References
-
- Arnold S. J., Robertson E. J. (2009). Making a commitment: cell lineage allocation and axis patterning in the early mouse embryo. Nat. Rev. Mol. Cell Biol. 10, 91-103 - PubMed
-
- Batlle-Morera L., Smith A., Nichols J. (2008). Parameters influencing derivation of embryonic stem cells from murine embryos. Genesis 46, 758-767 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
