

Nuclear lamins
- PMID: 20826548
- PMCID: PMC2964183
- DOI: 10.1101/cshperspect.a000547
Nuclear lamins
Abstract
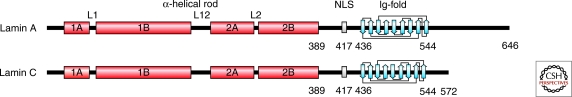
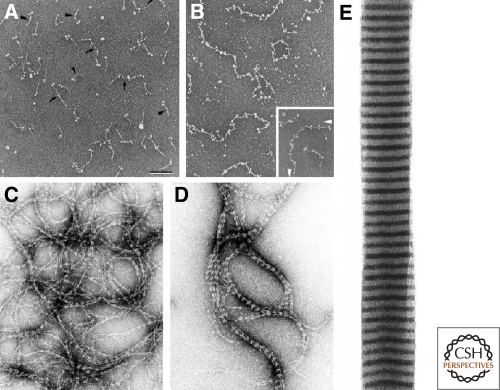
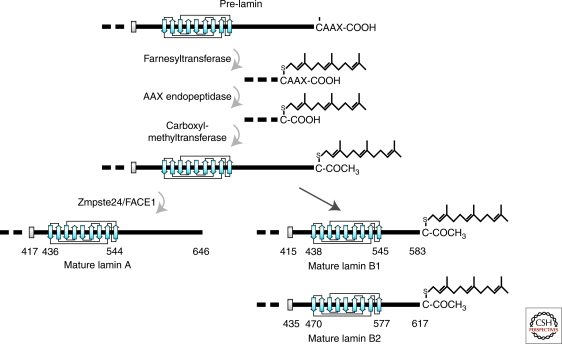
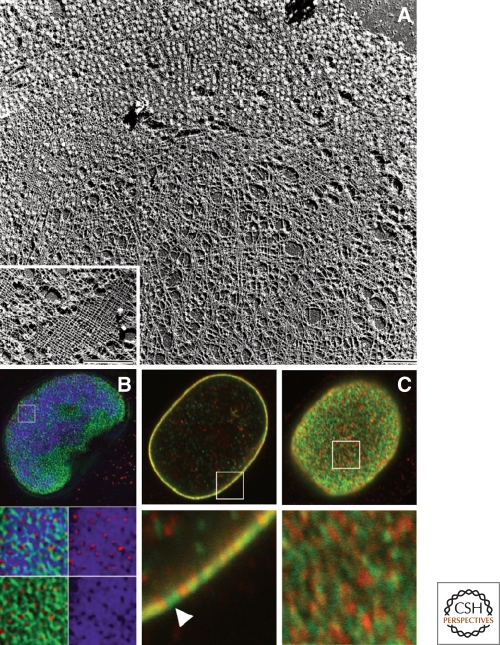
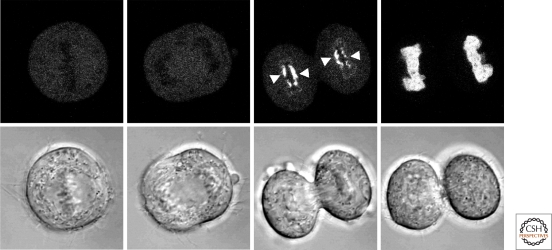
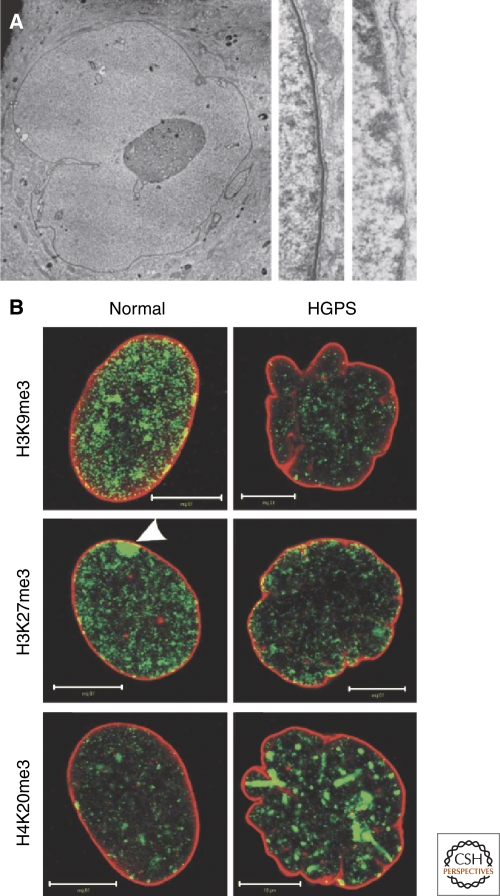
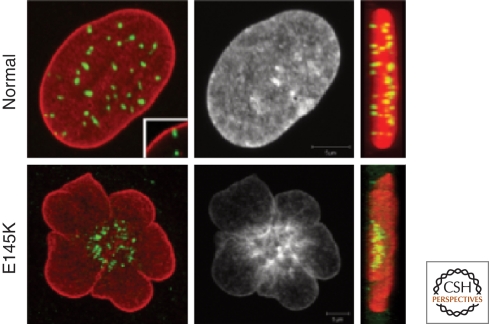
The nuclear lamins are type V intermediate filament proteins that are critically important for the structural properties of the nucleus. In addition, they are involved in the regulation of numerous nuclear processes, including DNA replication, transcription and chromatin organization. The developmentally regulated expression of lamins suggests that they are involved in cellular differentiation. Their assembly dynamic properties throughout the cell cycle, particularly in mitosis, are influenced by posttranslational modifications. Lamins may regulate nuclear functions by direct interactions with chromatin and determining the spatial organization of chromosomes within the nuclear space. They may also regulate chromatin functions by interacting with factors that epigenetically modify the chromatin or directly regulate replication or transcription.
Figures


References
-
- Adolph KW 1987. ADPribosylation of nuclear proteins labeled with [3H]adenosine: changes during the HeLa cycle. Biochim Biophys Acta 909: 222–230 - PubMed
-
- Aebi U, Cohn J, Buhle L, Gerace L 1986. The nuclear lamina is a meshwork of intermediate-type filaments. Nature 323: 560–564 - PubMed
-
- Akter R, Rivas D, Geneau G, Drissi H, Duque G 2009. Effect of lamin A/C knockdown on osteoblast differentiation and function. J Bone Miner Res 24: 283–293 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources