

A transmembrane amino acid in the GABAA receptor β2 subunit critical for the actions of alcohols and anesthetics
- PMID: 20826568
- PMCID: PMC2993559
- DOI: 10.1124/jpet.110.170472
A transmembrane amino acid in the GABAA receptor β2 subunit critical for the actions of alcohols and anesthetics
Abstract
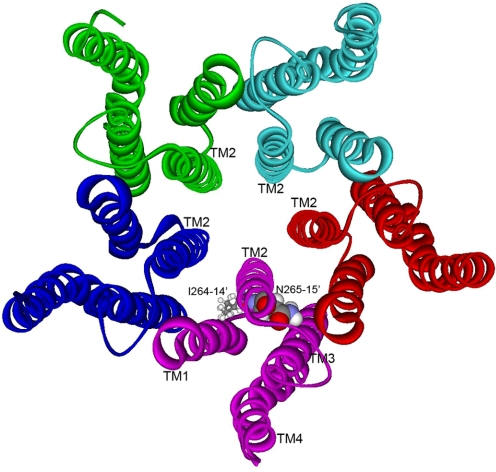
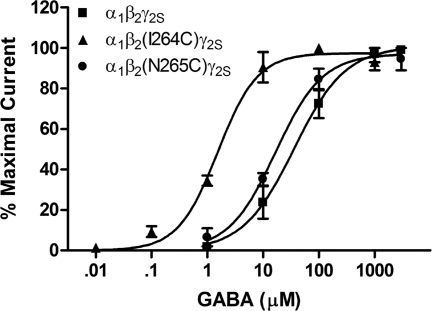
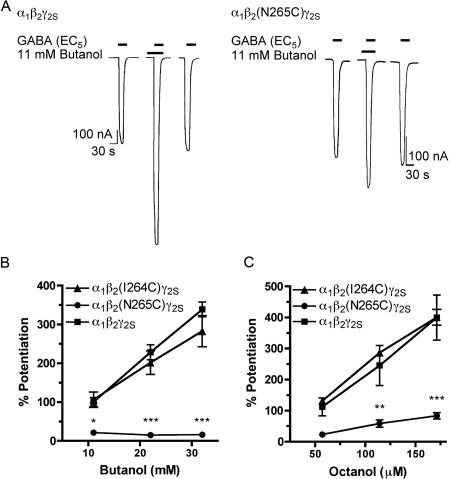
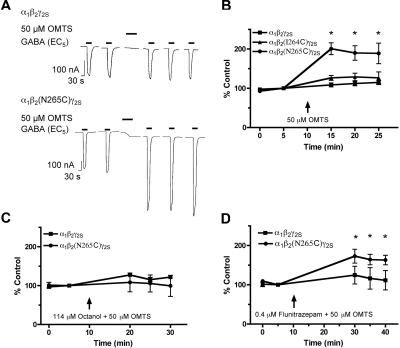
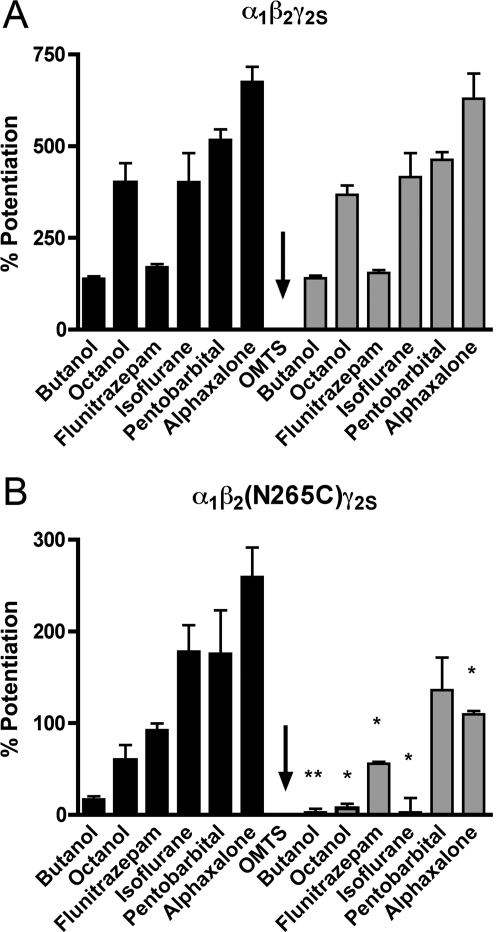
Alcohols and inhaled anesthetics enhance the function of GABA(A) receptors containing α, β, and γ subunits. Molecular analysis has focused on the role of the α subunits; however, there is evidence that the β subunits may also be important. The goal of our study was to determine whether Asn265, which is homologous to the site implicated in the α subunit (Ser270), contributes to an alcohol and volatile anesthetic binding site in the GABA(A) receptor β(2) subunit. We substituted cysteine for Asn265 and exposed the mutant to the sulfhydryl-specific reagent octyl methanethiosulfonate (OMTS). We used two-electrode voltage-clamp electrophysiology in Xenopus laevis oocytes and found that, after OMTS application, GABA-induced currents were irreversibly potentiated in mutant α(1)β(2)(N265C)γ(2S) receptors [but not α(1)β(2)(I264C)γ(2S)], presumably because of the covalent linking of octanethiol to the thiol group in the substituted cysteine. It is noteworthy that this effect was blocked when OMTS was applied in the presence of octanol. We found that potentiation by butanol, octanol, or isoflurane in the N265C mutant was nearly abolished after the application of OMTS, suggesting that an alcohol and volatile anesthetic binding site at position 265 of the β(2) subunit was irreversibly occupied by octanethiol and consequently prevented butanol or isoflurane from binding and producing their effects. OMTS did not affect modulation or direct activation by pentobarbital, but there was a partial reduction of allosteric modulation by flunitrazepam and alphaxalone in mutant α(1)β(2)(N265C)γ(2S) receptors after OMTS was applied. Our findings provide evidence that Asn265 may contribute to an alcohol and anesthetic binding site.
Figures
References
-
- Bali M, Akabas MH. (2004) Defining the propofol binding site location on the GABAA receptor. Mol Pharmacol 65:68–76 - PubMed
-
- Bocquet N, Nury H, Baaden M, Le Poupon C, Changeux JP, Delarue M, Corringer PJ. (2009) X-ray structure of a pentameric ligand-gated ion channel in an apparently open conformation. Nature 457:111–114 - PubMed
-
- Borghese EM, Henderson LA, Bleck Y, Trudell JR, Harris RA. (2003) Sites of excitatory and inhibitory actions of alcohols on neuronal alpha2beta4 nicotinic acetylcholine receptors. J Pharmacol Exp Ther 307:42–52 - PubMed
-
- Crawford DK, Trudell JR, Bertaccini EJ, Li K, Davies DL, Alkana RL. (2007) Evidence that ethanol acts on a target in Loop 2 of the extracellular domain of alpha1 glycine receptors. J Neurochem 102:2097–2109 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
