

Nanodomain control of exocytosis is responsible for the signaling capability of a retinal ribbon synapse
- PMID: 20826653
- PMCID: PMC2945284
- DOI: 10.1523/JNEUROSCI.1415-10.2010
Nanodomain control of exocytosis is responsible for the signaling capability of a retinal ribbon synapse
Abstract
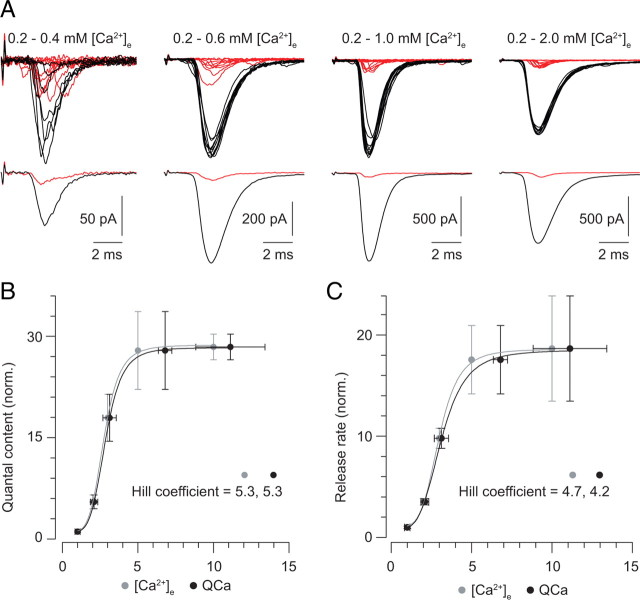
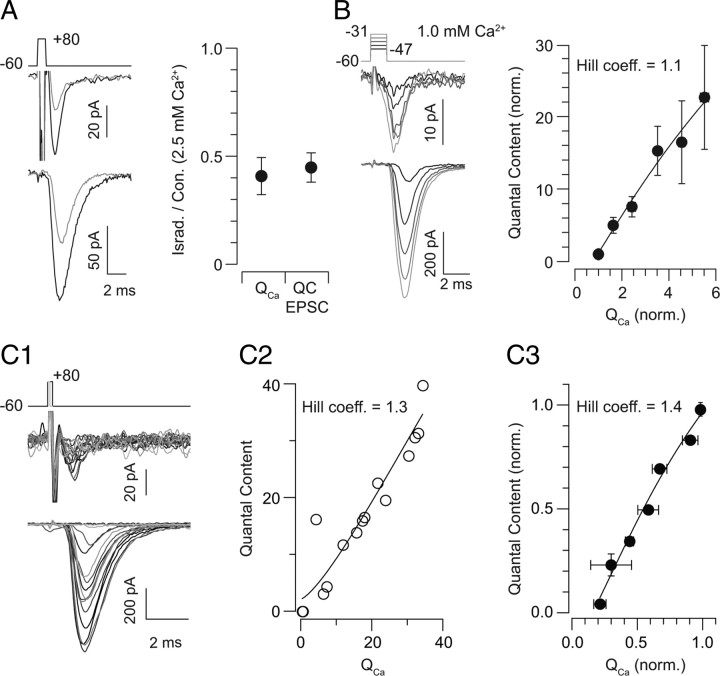
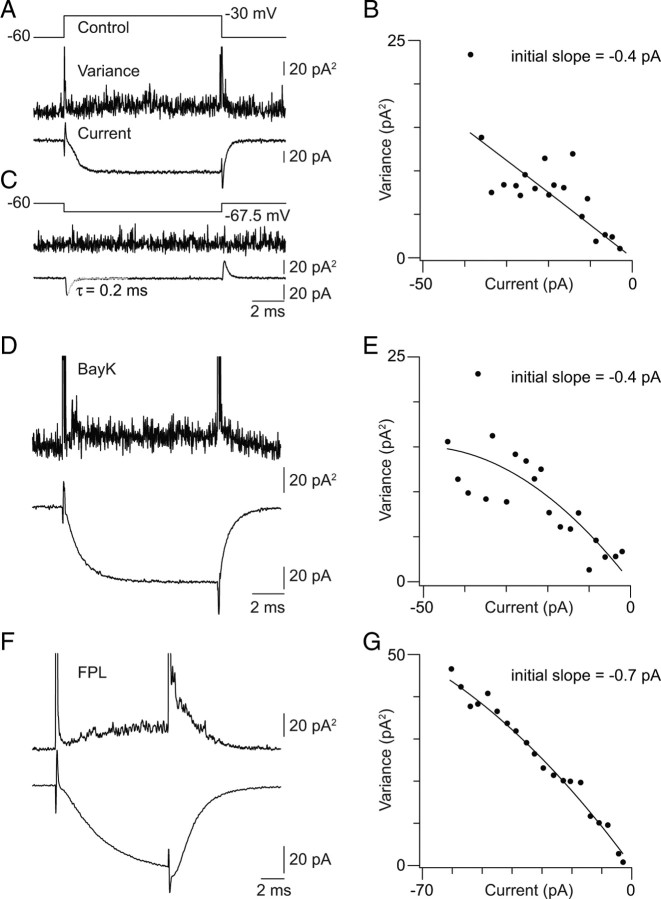
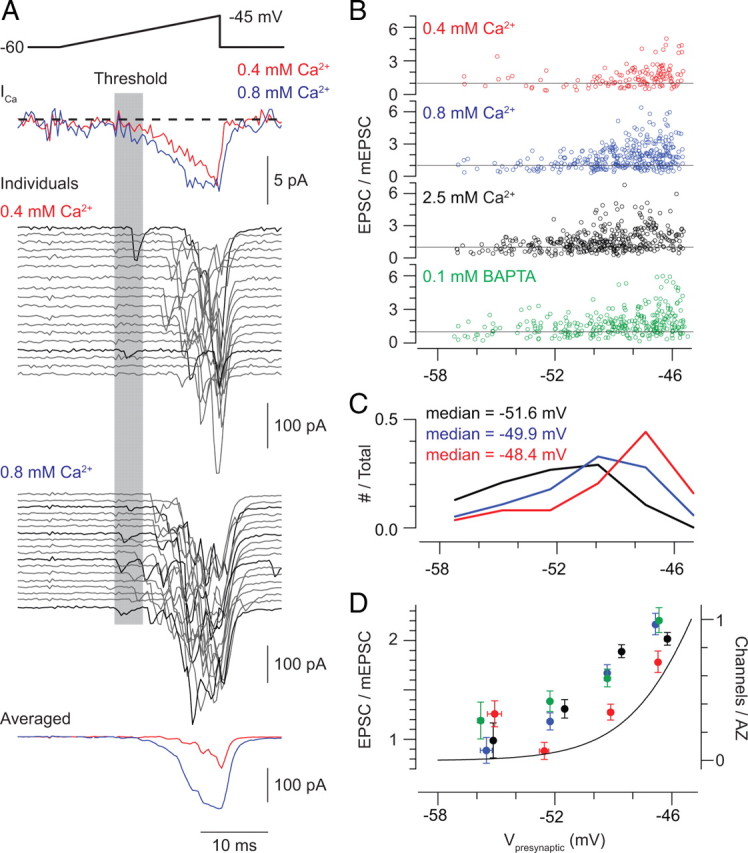
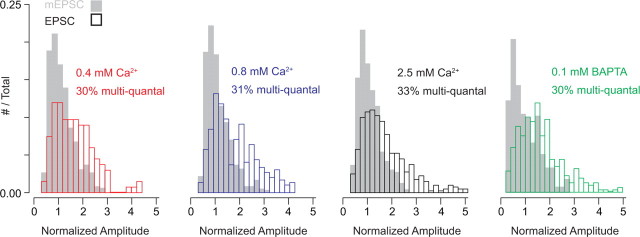
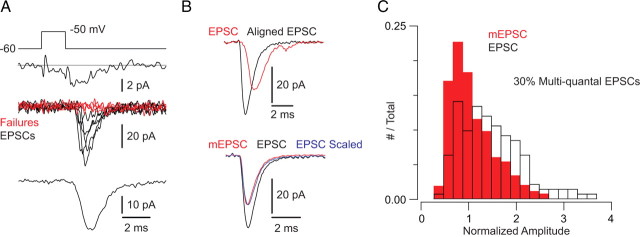
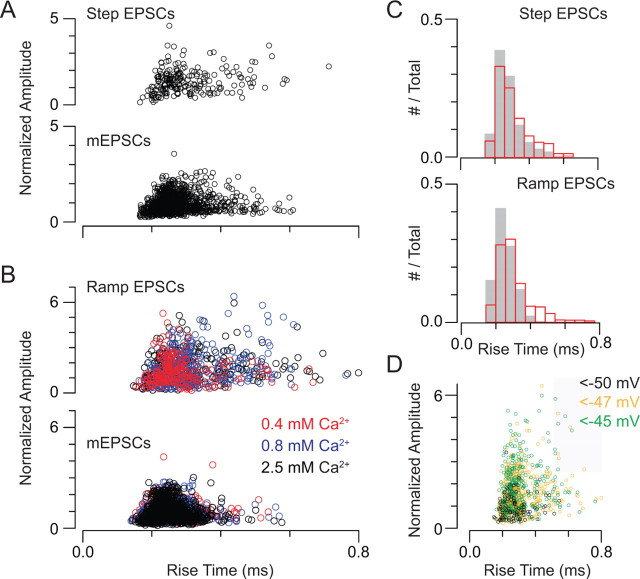
Primary sensory circuits encode both weak and intense stimuli reliably, requiring that their synapses signal over a wide dynamic range. In the retinal circuitry subserving night vision, processes intrinsic to the rod bipolar (RB) cell presynaptic active zone (AZ) permit the RB synapse to encode signals generated by the absorption of single photons as well as by more intense stimuli. In a study using an in vitro slice preparation of the mouse retina, we provide evidence that the location of Ca channels with low open probability within nanometers of the release sites is a critical determinant of the physiological behavior of the RB synapse. This gives rise to apparent one-to-one coupling between Ca channel opening and vesicle release, allowing presynaptic potential to be encoded linearly over a wide dynamic range. Further, it permits a transition from univesicular to multivesicular release (MVR) when two Ca channels/AZ open at potentials above the threshold for exocytosis. MVR permits small presynaptic voltage changes to elicit postsynaptic responses larger than quantal synaptic noise.
Figures
References
-
- Ames A, 3rd, Nesbett FB. In vitro retina as an experimental model of the central nervous system. J Neurochem. 1981;37:867–877. - PubMed
-
- Berjukow S, Marksteiner R, Gapp F, Sinnegger MJ, Hering S. Molecular mechanism of calcium channel block by isradipine. Role of a drug-induced inactivated channel conformation. J Biol Chem. 2000;275:22114–22120. - PubMed
-
- Berntson A, Taylor WR, Morgans CW. Molecular identity, synaptic localization, and physiology of calcium channels in retinal bipolar cells. J Neurosci Res. 2003;71:146–151. - PubMed
-
- Beutner D, Voets T, Neher E, Moser T. Calcium dependence of exocytosis and endocytosis at the cochlear inner hair cell afferent synapse. Neuron. 2001;29:681–690. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources