

Infectivity of hepatitis C virus is influenced by association with apolipoprotein E isoforms
- PMID: 20826689
- PMCID: PMC2977863
- DOI: 10.1128/JVI.01063-10
Infectivity of hepatitis C virus is influenced by association with apolipoprotein E isoforms
Abstract
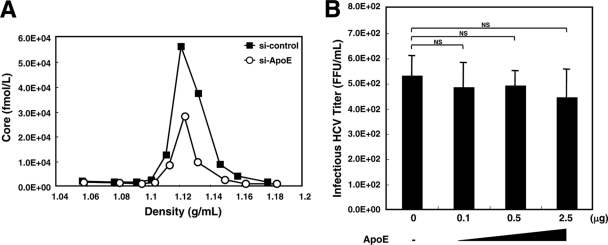
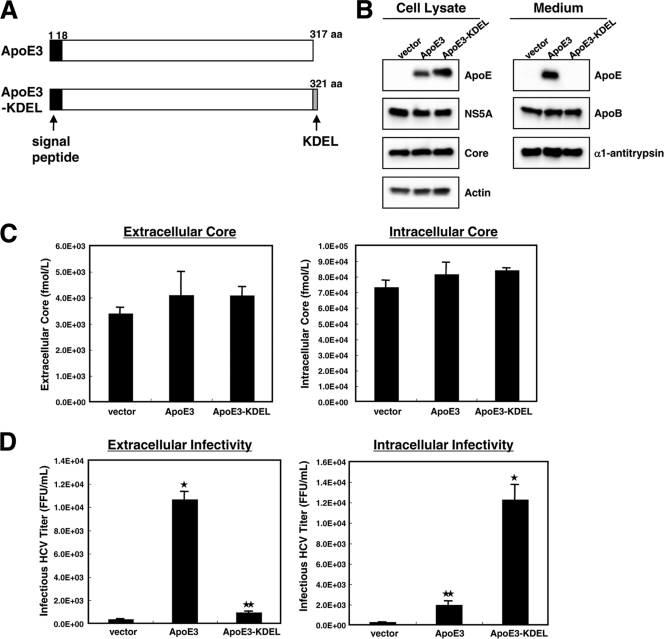
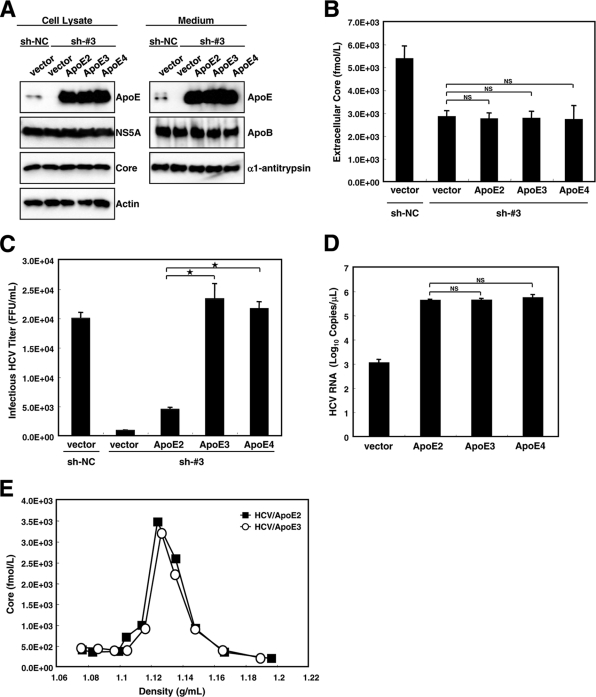
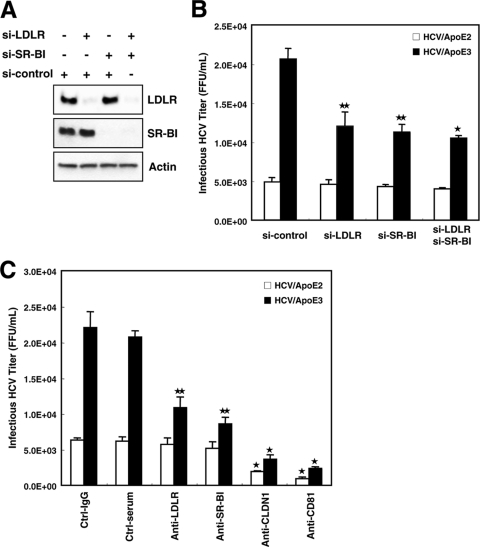
Hepatitis C virus (HCV) is a causative agent of chronic hepatitis, liver cirrhosis, and hepatocellular carcinoma. HCV in circulating blood associates with lipoproteins such as very low density lipoprotein (VLDL) and low-density lipoprotein (LDL). Although these associations suggest that lipoproteins are important for HCV infectivity, the roles of lipoproteins in HCV production and infectivity are not fully understood. To clarify the roles of lipoprotein in the HCV life cycle, we analyzed the effect of apolipoprotein E (ApoE), a component of lipoprotein, on virus production and infectivity. The production of infectious HCV was significantly reduced by the knockdown of ApoE. When an ApoE mutant that fails to be secreted into the culture medium was used, the amount of infectious HCV in the culture medium was dramatically reduced; the infectious HCV accumulated inside these cells, suggesting that infectious HCV must associate with ApoE prior to virus release. We performed rescue experiments in which ApoE isoforms were ectopically expressed in cells depleted of endogenous ApoE. The ectopic expression of the ApoE2 isoform, which has low affinity for the LDL receptor (LDLR), resulted in poor recovery of infectious HCV, whereas the expression of other isoforms, ApoE3 and ApoE4, rescued the production of infectious virus, raising it to an almost normal level. Furthermore, we found that the infectivity of HCV required both the LDLR and scavenger receptor class B, member I (SR-BI), ligands for ApoE. These findings indicate that ApoE is an essential apolipoprotein for HCV infectivity.
Figures
References
-
- Bartosch, B., A. Vitelli, C. Granier, C. Goujon, J. Dubuisson, S. Pascale, E. Scarselli, R. Cortese, A. Nicosia, and F. L. Cosset. 2003. Cell entry of hepatitis C virus requires a set of co-receptors that include the CD81 tetraspanin and the SR-B1 scavenger receptor. J. Biol. Chem. 278:41624-41630. - PubMed
-
- Benga, W. J., S. E. Krieger, M. Dimitrova, M. B. Zeisel, M. Parnot, J. Lupberger, E. Hildt, G. Luo, J. McLauchlan, T. F. Baumert, and C. Schuster. 2010. Apolipoprotein E interacts with hepatitis C virus nonstructural protein 5A and determines assembly of infectious particles. Hepatology 51:43-53. - PubMed
-
- Blasiole, D. A., R. A. Davis, and A. D. Attie. 2007. The physiological and molecular regulation of lipoprotein assembly and secretion. Mol. Biosyst. 3:608-619. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
