

Sustained glutamate receptor activation down-regulates GABAB receptors by shifting the balance from recycling to lysosomal degradation
- PMID: 20826795
- PMCID: PMC2975185
- DOI: 10.1074/jbc.M110.142406
Sustained glutamate receptor activation down-regulates GABAB receptors by shifting the balance from recycling to lysosomal degradation
Abstract
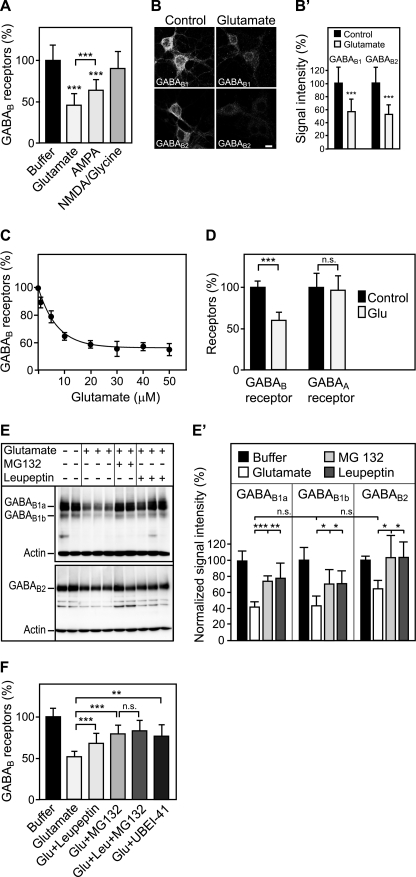
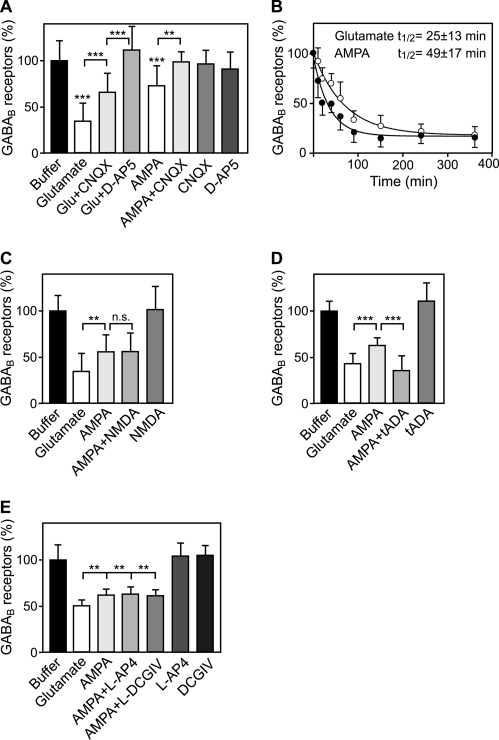
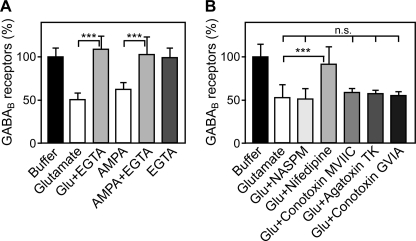
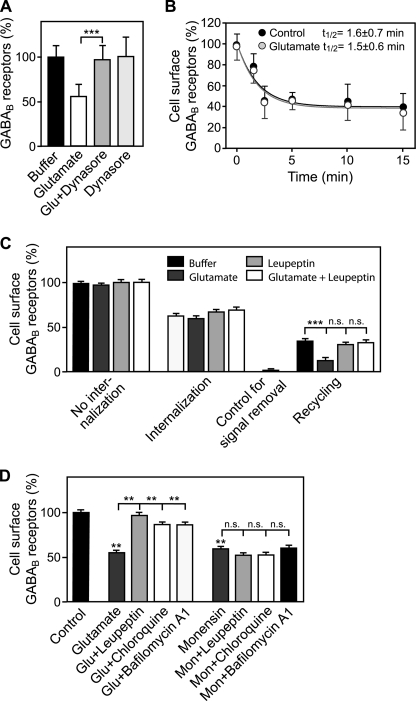
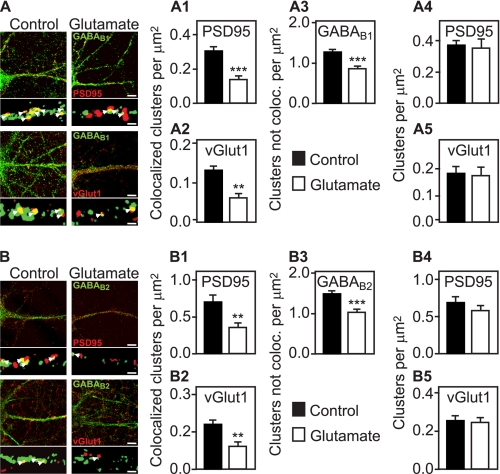
Metabotropic GABA(B) receptors are abundantly expressed at glutamatergic synapses where they control excitability of the synapse. Here, we tested the hypothesis that glutamatergic neurotransmission may regulate GABA(B) receptors. We found that application of glutamate to cultured cortical neurons led to rapid down-regulation of GABA(B) receptors via lysosomal degradation. This effect was mimicked by selective activation of AMPA receptors and further accelerated by coactivation of group I metabotropic glutamate receptors. Inhibition of NMDA receptors, blockade of L-type Ca(2+) channels, and removal of extracellular Ca(2+) prevented glutamate-induced down-regulation of GABA(B) receptors, indicating that Ca(2+) influx plays a critical role. We further established that glutamate-induced down-regulation depends on the internalization of GABA(B) receptors. Glutamate did not affect the rate of GABA(B) receptor endocytosis but led to reduced recycling of the receptors back to the plasma membrane. Blockade of lysosomal activity rescued receptor recycling, indicating that glutamate redirects GABA(B) receptors from the recycling to the degradation pathway. In conclusion, the data indicate that sustained activation of AMPA receptors down-regulates GABA(B) receptors by sorting endocytosed GABA(B) receptors preferentially to lysosomes for degradation on the expense of recycling. This mechanism may relieve glutamatergic synapses from GABA(B) receptor-mediated inhibition resulting in increased synaptic excitability.
Figures
References
-
- Bettler B., Kaupmann K., Mosbacher J., Gassmann M. (2004) Physiol. Rev. 84, 835–867 - PubMed
-
- Chen L., Boyes J., Yung W. H., Bolam J. P. (2004) J. Comp. Neurol. 474, 340–352 - PubMed
-
- Lacey C. J., Boyes J., Gerlach O., Chen L., Magill P. J., Bolam J. P. (2005) Neuroscience 136, 1083–1095 - PubMed
-
- Luján R., Shigemoto R., Kulik A., Juiz J. M. (2004) J. Comp. Neurol. 475, 36–46 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
