

Biochemical characterization of the transcriptional regulator BzdR from Azoarcus sp. CIB
- PMID: 20826820
- PMCID: PMC2975194
- DOI: 10.1074/jbc.M110.143503
Biochemical characterization of the transcriptional regulator BzdR from Azoarcus sp. CIB
Abstract
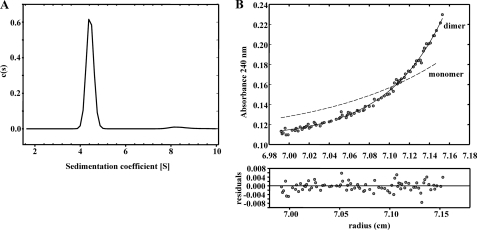
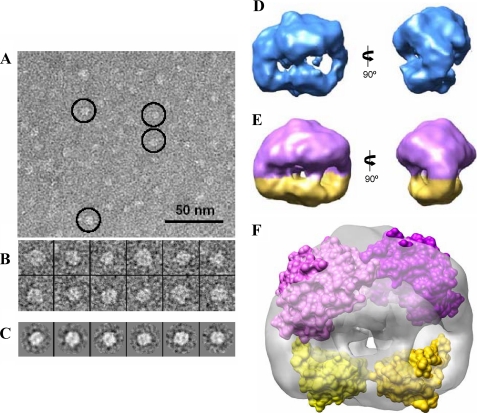
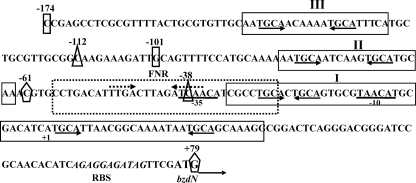
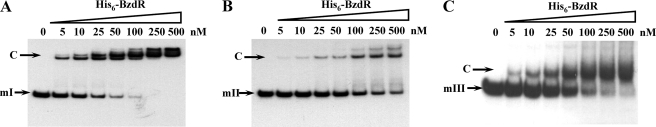
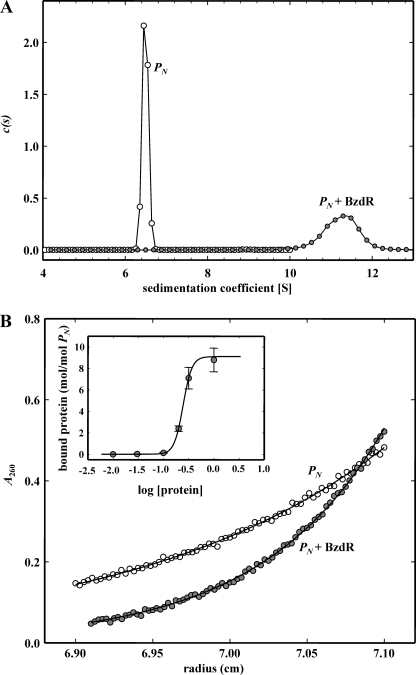
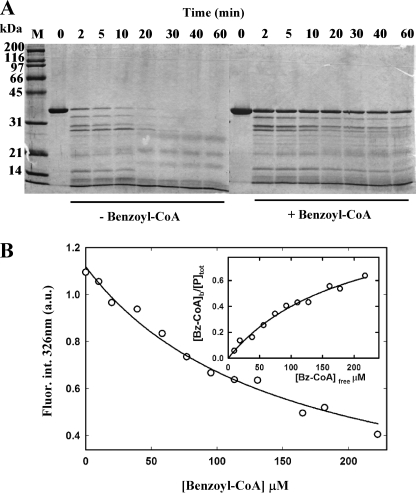
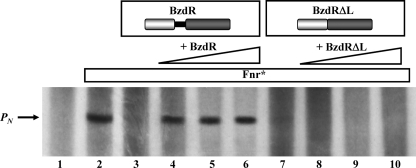
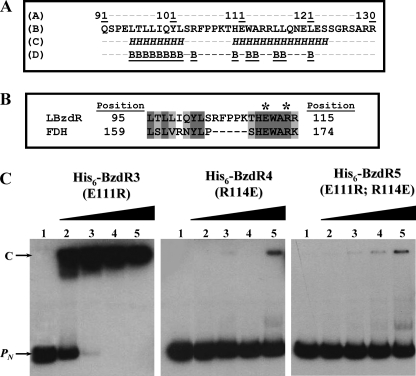
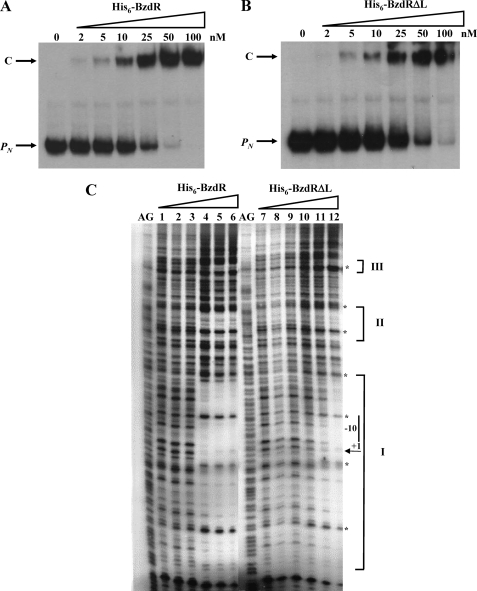
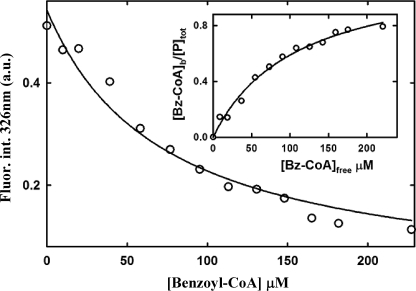
The BzdR transcriptional regulator that controls the P(N) promoter responsible for the anaerobic catabolism of benzoate in Azoarcus sp. CIB constitutes the prototype of a new subfamily of transcriptional regulators. Here, we provide some insights about the functional-structural relationships of the BzdR protein. Analytical ultracentrifugation studies revealed that BzdR is homodimeric in solution. An electron microscopy three-dimensional reconstruction of the BzdR dimer has been obtained, and the predicted structures of the respective N- and C-terminal domains of each BzdR monomer could be fitted into such a reconstruction. Gel retardation and ultracentrifugation experiments have shown that the binding of BzdR to its cognate promoter is cooperative. Different biochemical approaches revealed that the effector molecule benzoyl-CoA induces conformational changes in BzdR without affecting its oligomeric state. The BzdR-dependent inhibition of the P(N) promoter and its activation in the presence of benzoyl-CoA have been established by in vitro transcription assays. The monomeric BzdR4 and BzdR5 mutant regulators revealed that dimerization of BzdR is essential for DNA binding. Remarkably, a BzdRΔL protein lacking the linker region connecting the N- and C-terminal domains of BzdR is also dimeric and behaves as a super-repressor of the P(N) promoter. These data suggest that the linker region of BzdR is not essential for protein dimerization, but rather it is required to transfer the conformational changes induced by the benzoyl-CoA to the DNA binding domain leading to the release of the repressor. A model of action of the BzdR regulator has been proposed.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
