

The dithiol glutaredoxins of african trypanosomes have distinct roles and are closely linked to the unique trypanothione metabolism
- PMID: 20826822
- PMCID: PMC2966136
- DOI: 10.1074/jbc.M110.165860
The dithiol glutaredoxins of african trypanosomes have distinct roles and are closely linked to the unique trypanothione metabolism
Abstract
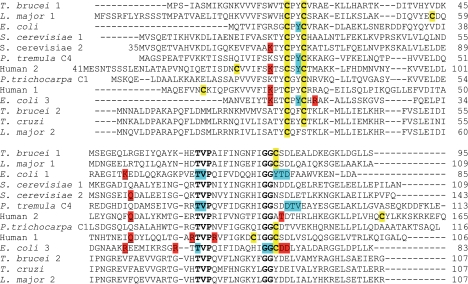
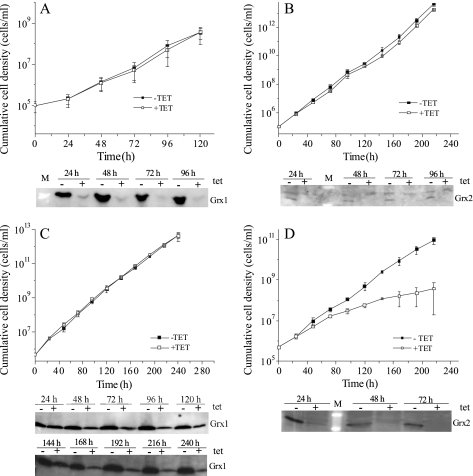
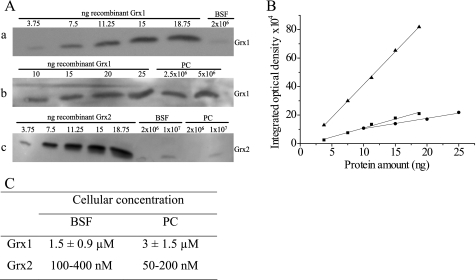
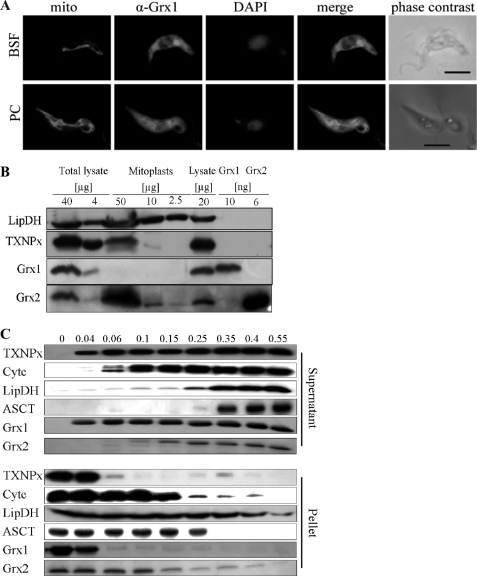
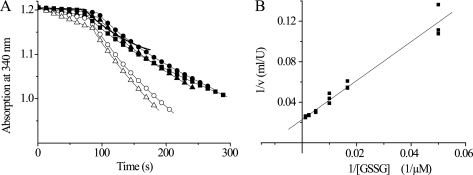
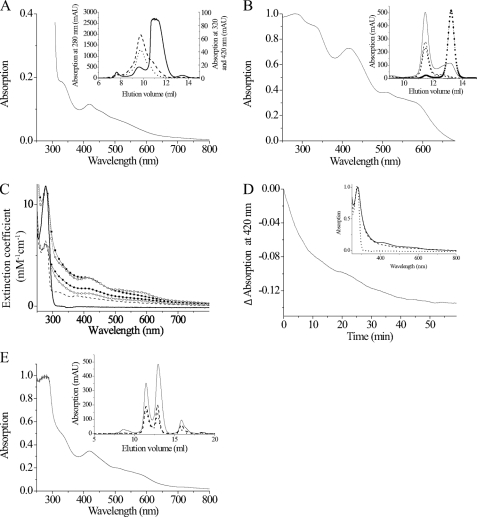
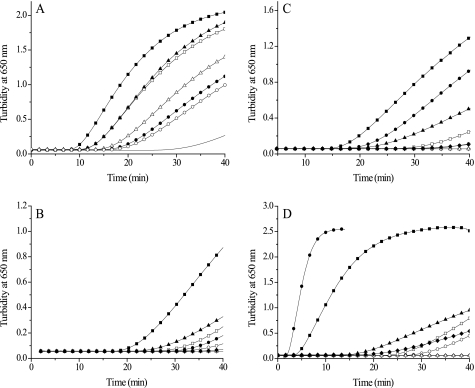
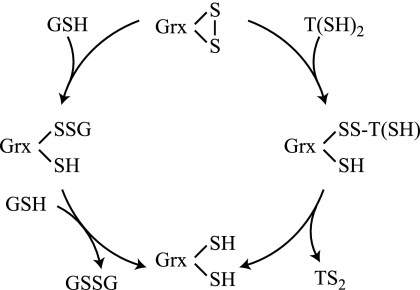
Trypanosoma brucei, the causative agent of African sleeping sickness, possesses two dithiol glutaredoxins (Grx1 and Grx2). Grx1 occurs in the cytosol and catalyzes protein deglutathionylations with k(cat)/K(m)-values of up to 2 × 10(5) M(-1) S(-1). It accelerates the reduction of ribonucleotide reductase by trypanothione although less efficiently than the parasite tryparedoxin and has low insulin disulfide reductase activity. Despite its classical CPYC active site, Grx1 forms dimeric iron-sulfur complexes with GSH, glutathionylspermidine, or trypanothione as non-protein ligands. Thus, contrary to the generally accepted assumption, replacement of the Pro is not a prerequisite for cluster formation. T. brucei Grx2 shows an unusual CQFC active site, and orthologues occur exclusively in trypanosomatids. Grx2 is enriched in mitoplasts, and fractionated digitonin lysis resulted in a co-elution with cytochrome c, suggesting localization in the mitochondrial intermembrane space. Grx2 catalyzes the reduction of insulin disulfide but not of ribonucleotide reductase and exerts deglutathionylation activity 10-fold lower than that of Grx1. RNA interference against Grx2 caused a growth retardation of procyclic cells consistent with an essential role. Grx1 and Grx2 are constitutively expressed with cellular concentrations of about 2 μM and 200 nM, respectively, in both the mammalian bloodstream and insect procyclic forms. Trypanothione reduces the disulfide form of both proteins with apparent rate constants that are 3 orders of magnitude higher than those with glutathione. Grx1 and, less efficiently, also Grx2 catalyze the reduction of GSSG by trypanothione. Thus, the Grxs play exclusive roles in the trypanothione-based thiol redox metabolism of African trypanosomes.
Figures
References
-
- Pai H. V., Starke D. W., Lesnefsky E. J., Hoppel C. L., Mieyal J. J. (2007) Antioxid. Redox Signal. 9, 2027–2033 - PubMed
-
- Lönn M. E., Hudemann C., Berndt C., Cherkasov V., Capani F., Holmgren A., Lillig C. H. (2008) Antioxid. Redox Signal. 10, 547–557 - PubMed
-
- Lillig C. H., Berndt C., Holmgren A. (2008) Biochim. Biophys. Acta 1780, 1304–1317 - PubMed
-
- Fernandes A. P., Holmgren A. (2004) Antioxid. Redox Signal. 6, 63–74 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
