

Astrocytes and pericytes differentially modulate blood-brain barrier characteristics during development and hypoxic insult
- PMID: 20827262
- PMCID: PMC3049523
- DOI: 10.1038/jcbfm.2010.148
Astrocytes and pericytes differentially modulate blood-brain barrier characteristics during development and hypoxic insult
Abstract
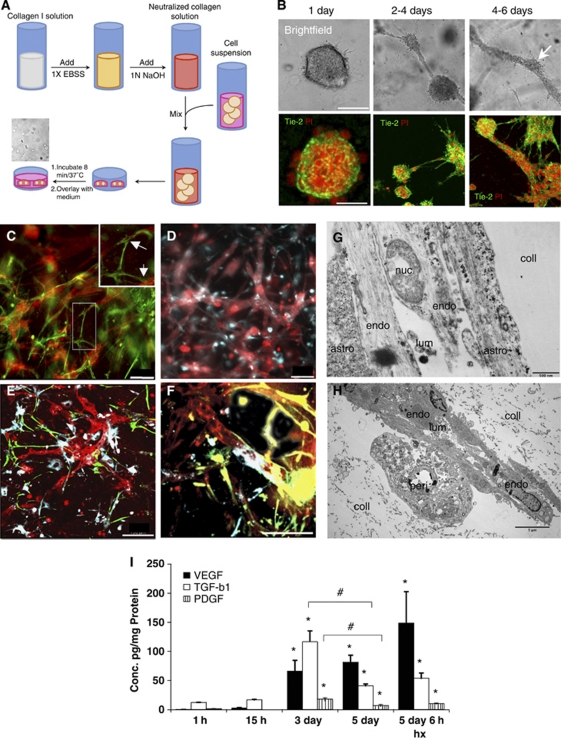
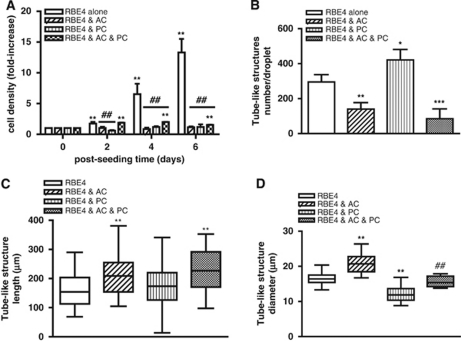
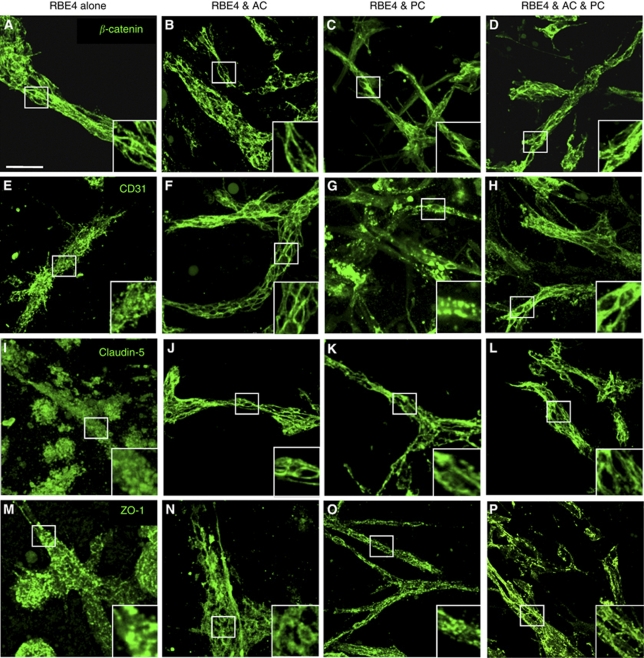
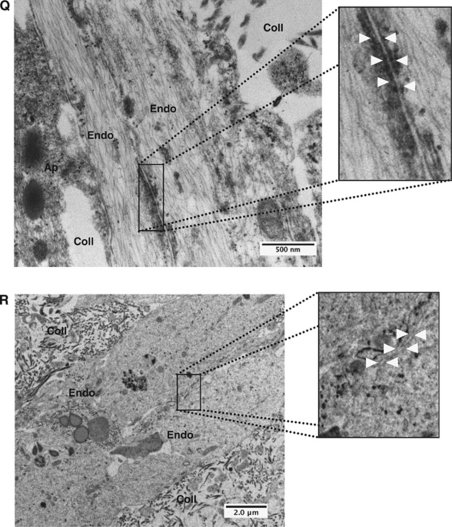
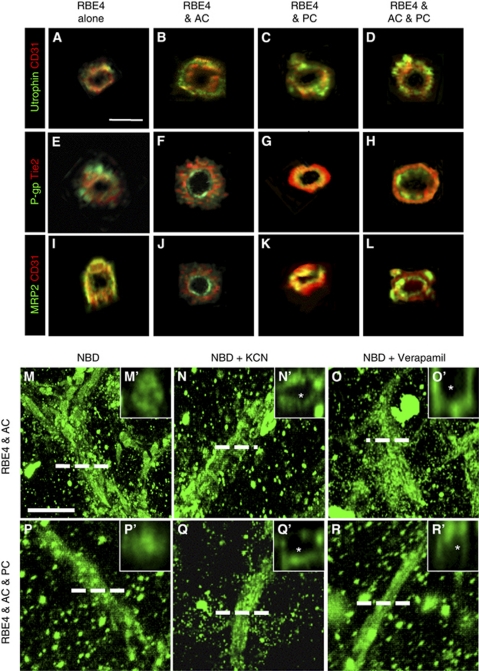
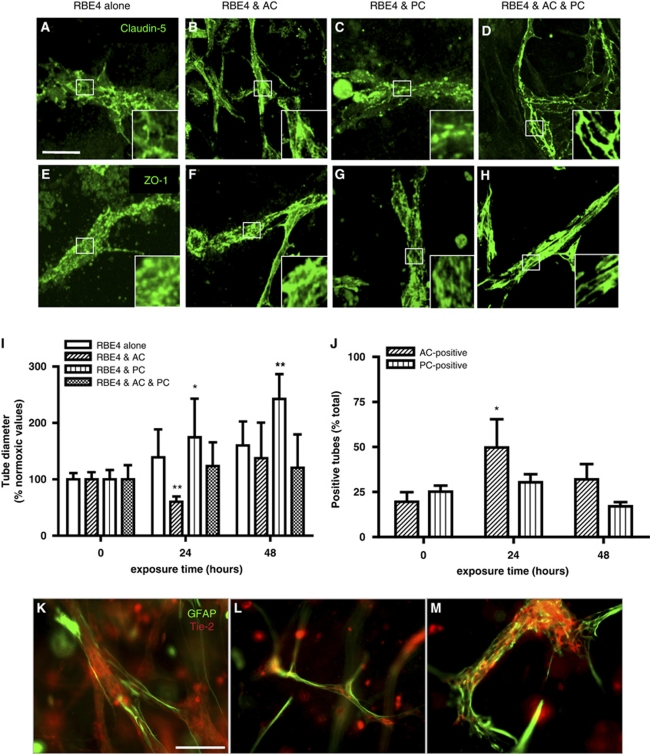
Understanding regulation of blood-brain barrier (BBB) is crucial to reduce/prevent its disruption during injury. As high brain complexity makes interpretation of in vivo data challenging, BBB studies are frequently performed using simplified in vitro models. However, many models fail to address the three-dimensional (3D) cellular interactions that occur in vivo, an important feature that may explain discrepancies in translation of in vitro data to the in vivo situation. We have designed and characterized an innovative 3D model that reproduces morphological and functional characteristics of the BBB in vivo and used it to investigate cellular interactions and contribution of astrocytes and pericytes to BBB development. Our model shows that both astrocytes and pericytes significantly suppress endothelial proliferation. In contrast, differential effects on tubulogenesis were observed with astrocytes reducing the number of tubes formed but increasing diameters and length, whereas pericytes had the opposite effect. Pericytes also induce proper localization of barrier proteins, lumen polarization, and functional activity of ATP-binding cassette (ABC) transporters similar to astrocytes, but the presence of both cells is required to maintain optimal barrier characteristics during hypoxic exposure. This model is simple, dynamic, and convenient to study many aspects of BBB function and represents an exciting new tool to address open questions of BBB regulation.
Figures
References
-
- Al Ahmad A, Gassmann M, Ogunshola OO. Maintaining blood-brain barrier integrity: pericytes perform better than astrocytes during prolonged oxygen deprivation. J Cell Physiol. 2009;218:612–622. - PubMed
-
- Baldessari D, Mione M. How to create the vascular tree? (Latest) help from the zebrafish. Pharmacol Ther. 2008;118:206–230. - PubMed
-
- Banks WA, Ercal N, Price TO. The blood-brain barrier in neuroAIDS. Curr HIV Res. 2006;4:259–266. - PubMed
-
- Bauer B, Hartz AM, Fricker G, Miller DS. Pregnane X receptor up-regulation of P-glycoprotein expression and transport function at the blood-brain barrier. Mol Pharmacol. 2004;66:413–419. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
