

Lazarus1, a DUF300 protein, contributes to programmed cell death associated with Arabidopsis acd11 and the hypersensitive response
- PMID: 20830211
- PMCID: PMC2935358
- DOI: 10.1371/journal.pone.0012586
Lazarus1, a DUF300 protein, contributes to programmed cell death associated with Arabidopsis acd11 and the hypersensitive response
Abstract
Background: Programmed cell death (PCD) is a necessary part of the life of multi-cellular organisms. A type of plant PCD is the defensive hypersensitive response (HR) elicited via recognition of a pathogen by host resistance (R) proteins. The lethal, recessive accelerated cell death 11 (acd11) mutant exhibits HR-like accelerated cell death, and cell death execution in acd11 shares genetic requirements for HR execution triggered by one subclass of R proteins.
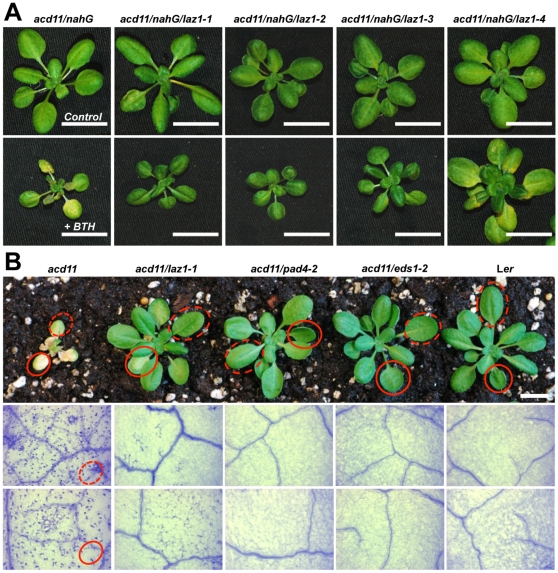
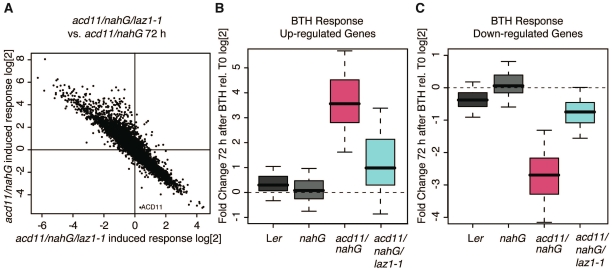
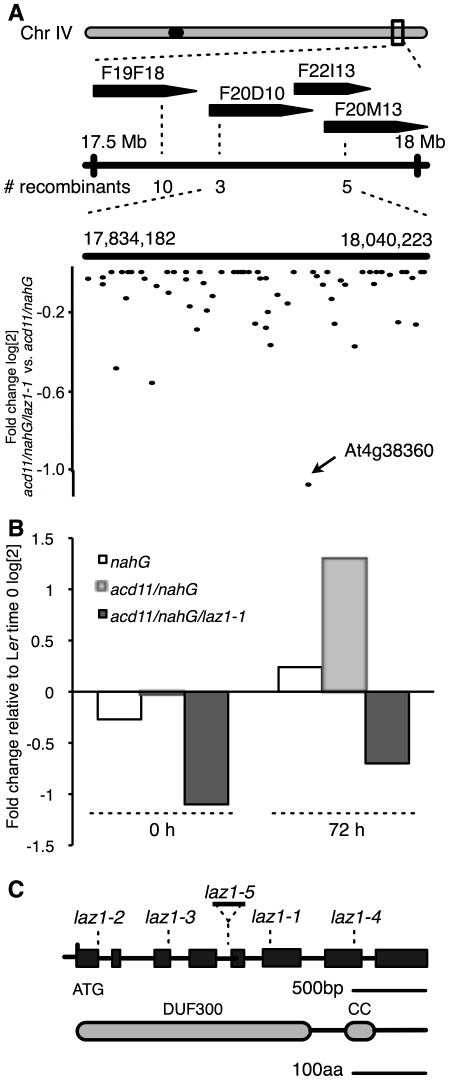
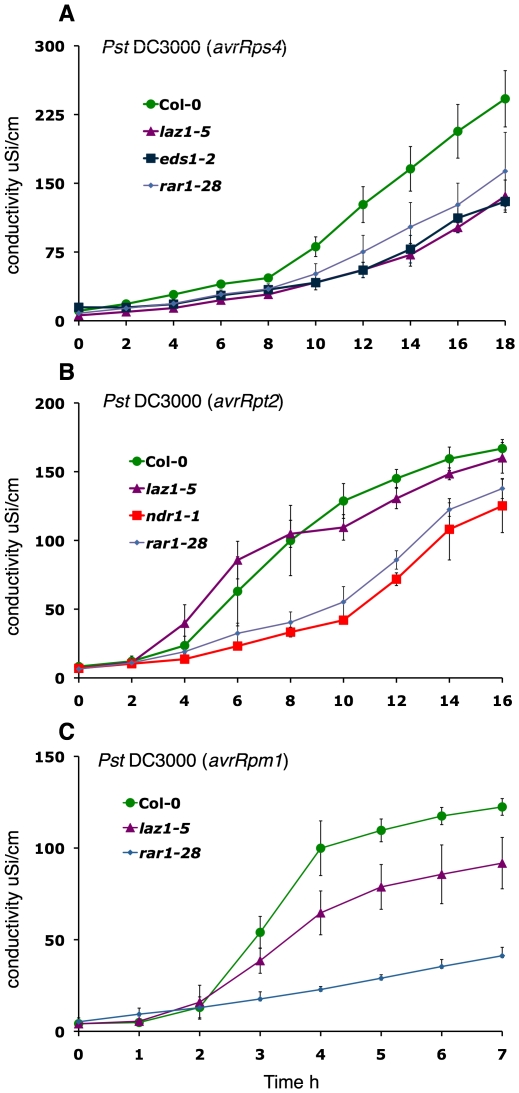
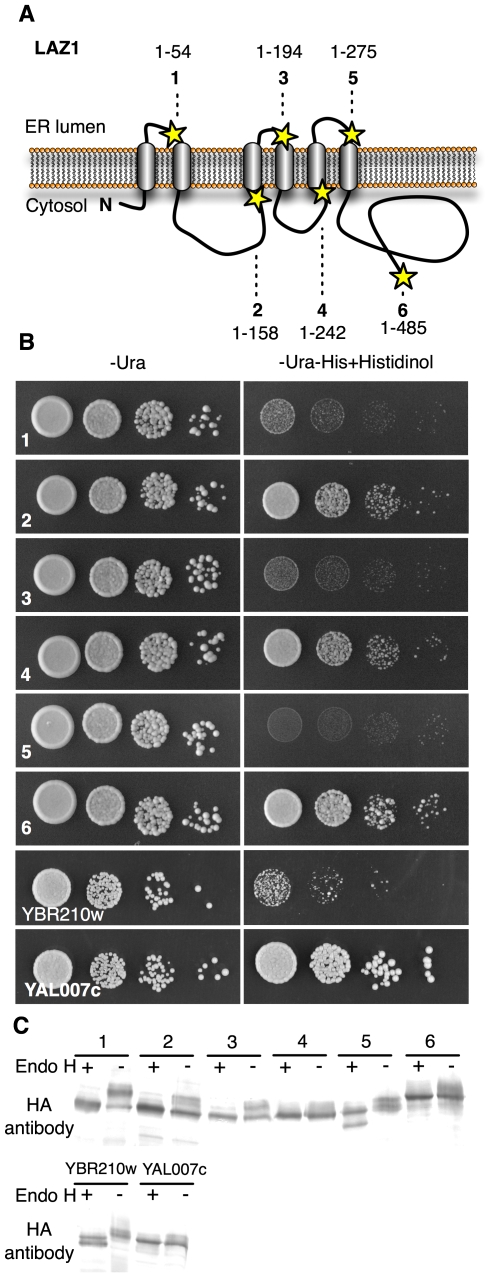
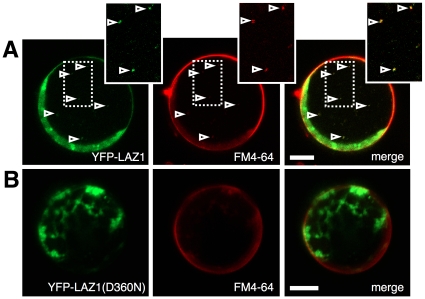
Methodology/principal findings: To identify genes required for this PCD pathway, we conducted a genetic screen for suppressors of acd11, here called lazarus (laz) mutants. In addition to known suppressors of R protein-mediated HR, we isolated 13 novel complementation groups of dominant and recessive laz mutants. Here we describe laz1, which encodes a protein with a domain of unknown function (DUF300), and demonstrate that LAZ1 contributes to HR PCD conditioned by the Toll/interleukin-1 (TIR)-type R protein RPS4 and by the coiled-coil (CC)-type R protein RPM1. Using a yeast-based topology assay, we also provide evidence that LAZ1 is a six transmembrane protein with structural similarities to the human tumor suppressor TMEM34. Finally, we demonstrate by transient expression of reporter fusions in protoplasts that localization of LAZ1 is distributed between the cytosol, the plasma membrane and FM4-64 stained vesicles.
Conclusions/significance: Our findings indicate that LAZ1 functions as a regulator or effector of plant PCD associated with the HR, in addition to its role in acd11-related death. Furthermore, the similar topology of a plant and human DUF300 proteins suggests similar functions in PCD across the eukaryotic kingdoms, although a direct role for TMEM34 in cell death control remains to be established. Finally, the subcellular localization pattern of LAZ1 suggests that it may have transport functions for yet unknown, death-related signaling molecules at the plasma membrane and/or endosomal compartments. In summary, our results validate the utility of the large-scale suppressor screen to identify novel components with functions in plant PCD, which may also have implications for deciphering cell death mechanisms in other organisms.
Conflict of interest statement
Figures
Similar articles
-
The LSD1-interacting protein GILP is a LITAF domain protein that negatively regulates hypersensitive cell death in Arabidopsis.PLoS One. 2011 Apr 19;6(4):e18750. doi: 10.1371/journal.pone.0018750. PLoS One. 2011. PMID: 21526181 Free PMC article.
-
Knockout of Arabidopsis accelerated-cell-death11 encoding a sphingosine transfer protein causes activation of programmed cell death and defense.Genes Dev. 2002 Feb 15;16(4):490-502. doi: 10.1101/gad.218202. Genes Dev. 2002. PMID: 11850411 Free PMC article.
-
Retromer contributes to immunity-associated cell death in Arabidopsis.Plant Cell. 2015 Feb;27(2):463-79. doi: 10.1105/tpc.114.132043. Epub 2015 Feb 13. Plant Cell. 2015. PMID: 25681156 Free PMC article.
-
The Evolutionary Origins of Programmed Cell Death Signaling.Cold Spring Harb Perspect Biol. 2020 Sep 1;12(9):a036442. doi: 10.1101/cshperspect.a036442. Cold Spring Harb Perspect Biol. 2020. PMID: 31818855 Free PMC article. Review.
-
Dynamic intracellular reorganization of cytoskeletons and the vacuole in defense responses and hypersensitive cell death in plants.J Plant Res. 2011 May;124(3):315-24. doi: 10.1007/s10265-011-0408-z. Epub 2011 Mar 16. J Plant Res. 2011. PMID: 21409543 Review.
Cited by
-
Analysis of the 5' Untranslated Region Length-Dependent Control of Gene Expression in Maize: A Case Study with the ZmLAZ1 Gene Family.Genes (Basel). 2024 Jul 29;15(8):994. doi: 10.3390/genes15080994. Genes (Basel). 2024. PMID: 39202355 Free PMC article.
-
The LSD1-interacting protein GILP is a LITAF domain protein that negatively regulates hypersensitive cell death in Arabidopsis.PLoS One. 2011 Apr 19;6(4):e18750. doi: 10.1371/journal.pone.0018750. PLoS One. 2011. PMID: 21526181 Free PMC article.
-
Genome-wide analysis of the LAZ1 gene family in Gossypium hirsutum.Mol Biol Rep. 2023 Nov;50(11):9273-9282. doi: 10.1007/s11033-023-08788-5. Epub 2023 Oct 9. Mol Biol Rep. 2023. PMID: 37812351
-
Sphingolipids and plant defense/disease: the "death" connection and beyond.Front Plant Sci. 2012 Apr 10;3:68. doi: 10.3389/fpls.2012.00068. eCollection 2012. Front Plant Sci. 2012. PMID: 22639658 Free PMC article.
-
BASIDIN as a New Protein Effector of the Phytopathogen Causing Witche's Broom Disease in Cocoa.Int J Mol Sci. 2023 Jul 20;24(14):11714. doi: 10.3390/ijms241411714. Int J Mol Sci. 2023. PMID: 37511472 Free PMC article.
References
-
- Baehrecke EH. How death shapes life during development. Nat Rev Mol Cell Biol. 2002;3:779–787. - PubMed
-
- Greenberg JT, Yao N. The role and regulation of programmed cell death in plant-pathogen interactions. Cell Microbiol. 2004;6:201–211. - PubMed
-
- Jones JD, Dangl JL. The plant immune system. Nature. 2006;444:323–329. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
