

Sexual dimorphism in the fly brain
- PMID: 20832311
- PMCID: PMC2957842
- DOI: 10.1016/j.cub.2010.07.045
Sexual dimorphism in the fly brain
Abstract
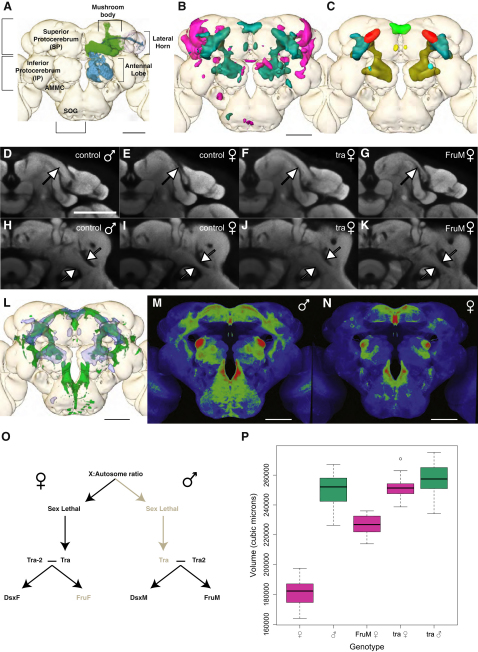
Background: Sex-specific behavior may originate from differences in brain structure or function. In Drosophila, the action of the male-specific isoform of fruitless in about 2000 neurons appears to be necessary and sufficient for many aspects of male courtship behavior. Initial work found limited evidence for anatomical dimorphism in these fru+ neurons. Subsequently, three discrete anatomical differences in central brain fru+ neurons have been reported, but the global organization of sex differences in wiring is unclear.
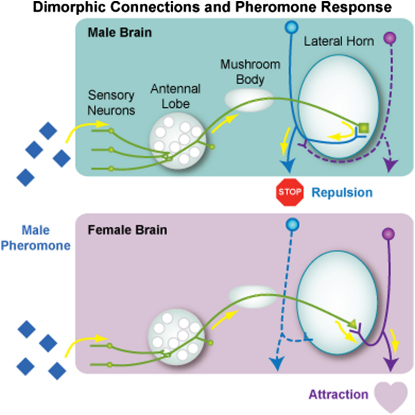
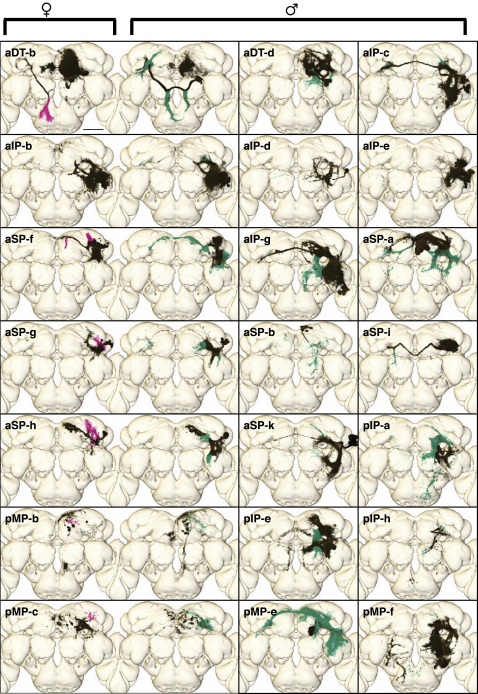
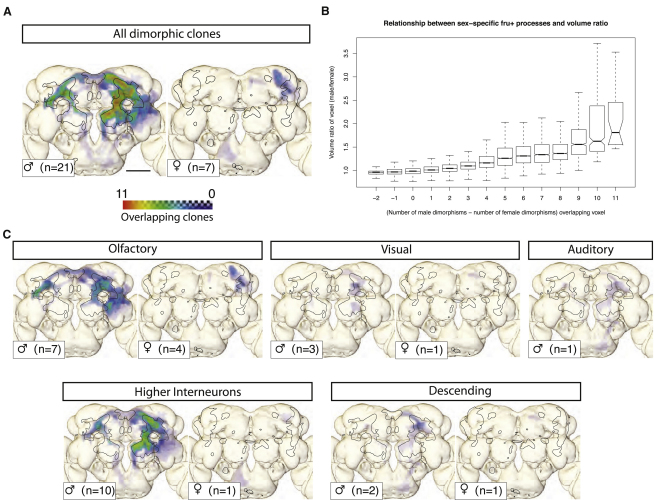
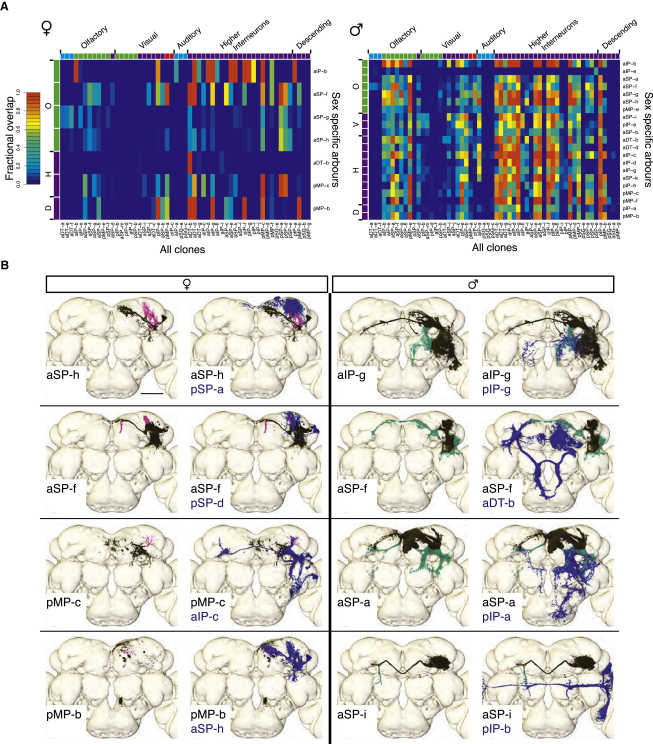
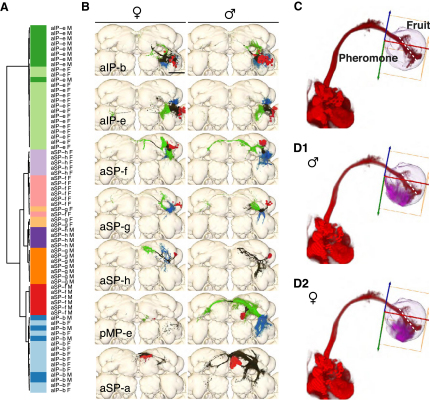
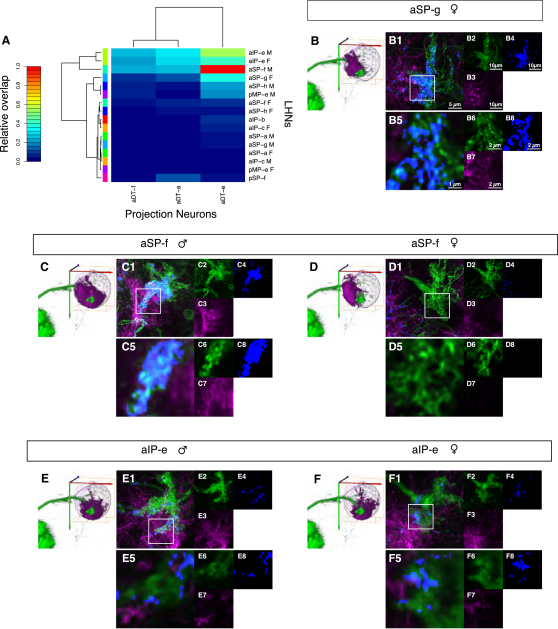
Results: A global search for structural differences in the Drosophila brain identified large volumetric differences between males and females, mostly in higher brain centers. In parallel, saturating clonal analysis of fru+ neurons using mosaic analysis with a repressible cell marker identified 62 neuroblast lineages that generate fru+ neurons in the brain. Coregistering images from male and female brains identified 19 new dimorphisms in males; these are highly concentrated in male-enlarged higher brain centers. Seven dimorphic lineages also had female-specific arbors. In addition, at least 5 of 51 fru+ lineages in the nerve cord are dimorphic. We use these data to predict >700 potential sites of dimorphic neural connectivity. These are particularly enriched in third-order olfactory neurons of the lateral horn, where we provide strong evidence for dimorphic anatomical connections by labeling partner neurons in different colors in the same brain.
Conclusion: Our analysis reveals substantial differences in wiring and gross anatomy between male and female fly brains. Reciprocal connection differences in the lateral horn offer a plausible explanation for opposing responses to sex pheromones in male and female flies.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
References
-
- Hall J.C. The mating of a fly. Science. 1994;264:1702–1714. - PubMed
-
- Baker B.S., Taylor B.J., Hall J.C. Are complex behaviors specified by dedicated regulatory genes? Reasoning from Drosophila. Cell. 2001;105:13–24. - PubMed
-
- Billeter J.C., Rideout E.J., Dornan A.J., Goodwin S.F. Control of male sexual behavior in Drosophila by the sex determination pathway. Curr. Biol. 2006;16:R766–R776. - PubMed
-
- Hotta Y., Benzer S. Mapping of behaviour in Drosophila mosaics. Nature. 1972;240:527–535. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
