

Comprehensive, fine-scale dissection of homologous recombination outcomes at a hot spot in mouse meiosis
- PMID: 20832722
- PMCID: PMC3196603
- DOI: 10.1016/j.molcel.2010.08.017
Comprehensive, fine-scale dissection of homologous recombination outcomes at a hot spot in mouse meiosis
Abstract
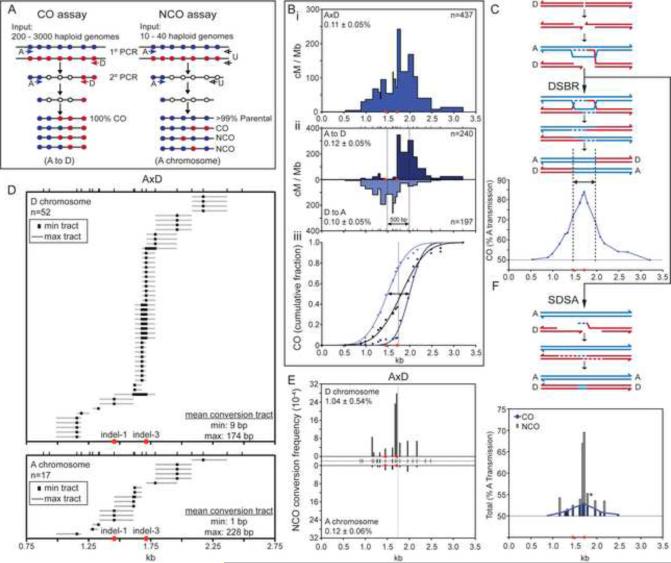
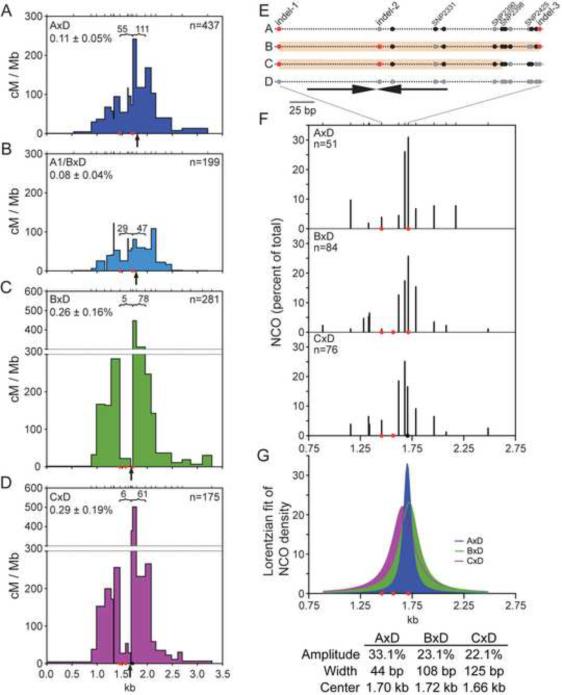
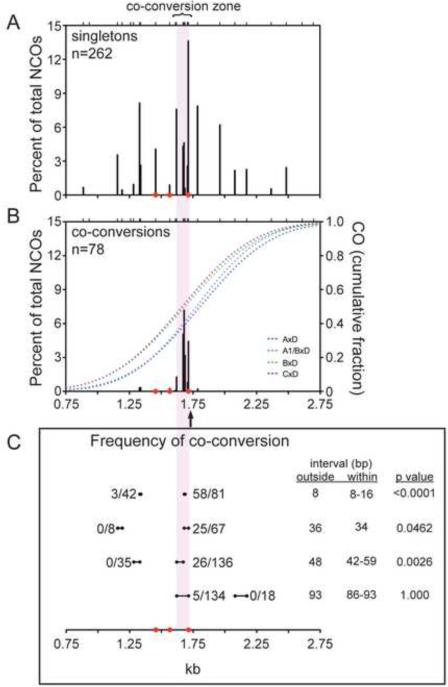
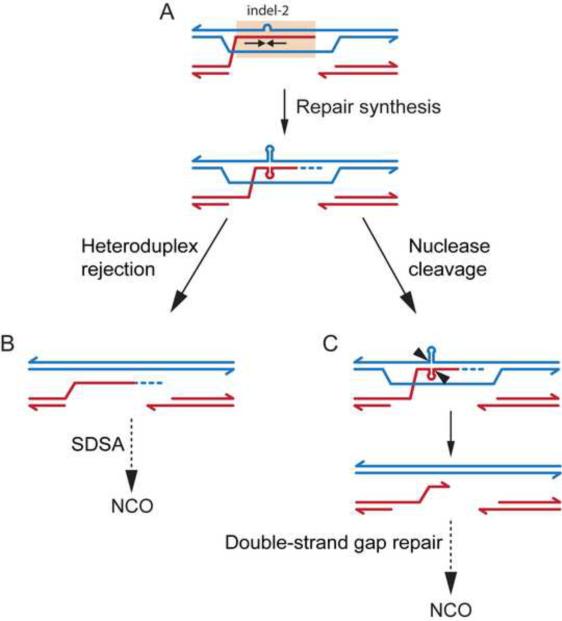
In mammalian meiosis, only a small fraction of programmed DNA double-strand breaks are repaired as interhomolog crossovers (COs). To analyze another product of meiotic recombination, interhomolog noncrossovers (NCOs), we performed high-resolution mapping of recombination events at an intensely active mouse hot spot in F1 hybrids derived from inbred mouse strains. We provide direct evidence that the vast majority of repair events are interhomolog NCOs, consistent with models in which frequent interhomolog interactions promote accurate chromosome pairing. NCOs peaked at the center of the hot spot but were also broadly distributed throughout. In some hybrid strains, localized zones within the hot spot were highly refractory to COs and showed elevated frequency of coconversion of adjacent polymorphisms in NCOs, raising the possibility of double-strand gap repair. Transmission distortion was observed in one hybrid, with NCOs providing a significant contribution. Thus, NCO recombination events play a substantial role in mammalian meiosis and genome evolution.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures

Similar articles
-
Local and sex-specific biases in crossover vs. noncrossover outcomes at meiotic recombination hot spots in mice.Genes Dev. 2015 Aug 15;29(16):1721-33. doi: 10.1101/gad.265561.115. Epub 2015 Aug 6. Genes Dev. 2015. PMID: 26251527 Free PMC article.
-
Elevated Rad53 kinase activity influences formation and interhomolog repair of meiotic DNA double-strand breaks in budding yeast.Biochem Biophys Res Commun. 2013 Nov 22;441(3):593-9. doi: 10.1016/j.bbrc.2013.10.111. Epub 2013 Oct 30. Biochem Biophys Res Commun. 2013. PMID: 24183725
-
The Hop2 protein has a direct role in promoting interhomolog interactions during mouse meiosis.Dev Cell. 2003 Dec;5(6):927-36. doi: 10.1016/s1534-5807(03)00369-1. Dev Cell. 2003. PMID: 14667414
-
The meiotic-specific Mek1 kinase in budding yeast regulates interhomolog recombination and coordinates meiotic progression with double-strand break repair.Curr Genet. 2019 Jun;65(3):631-641. doi: 10.1007/s00294-019-00937-3. Epub 2019 Jan 22. Curr Genet. 2019. PMID: 30671596 Free PMC article. Review.
-
Preaching about the converted: how meiotic gene conversion influences genomic diversity.Ann N Y Acad Sci. 2012 Sep;1267:95-102. doi: 10.1111/j.1749-6632.2012.06595.x. Ann N Y Acad Sci. 2012. PMID: 22954222 Free PMC article. Review.
Cited by
-
Initiation of meiotic homologous recombination: flexibility, impact of histone modifications, and chromatin remodeling.Cold Spring Harb Perspect Biol. 2015 May 1;7(5):a016527. doi: 10.1101/cshperspect.a016527. Cold Spring Harb Perspect Biol. 2015. PMID: 25934010 Free PMC article. Review.
-
Age-Dependent Alterations in Meiotic Recombination Cause Chromosome Segregation Errors in Spermatocytes.Cell. 2017 Oct 19;171(3):601-614.e13. doi: 10.1016/j.cell.2017.08.042. Epub 2017 Sep 21. Cell. 2017. PMID: 28942922 Free PMC article.
-
Interhomolog polymorphism shapes meiotic crossover within the Arabidopsis RAC1 and RPP13 disease resistance genes.PLoS Genet. 2018 Dec 13;14(12):e1007843. doi: 10.1371/journal.pgen.1007843. eCollection 2018 Dec. PLoS Genet. 2018. PMID: 30543623 Free PMC article.
-
Enhancing local meiotic crossovers in Arabidopsis and maize through juxtaposition of heterozygous and homozygous regions.Nat Plants. 2025 Sep 2. doi: 10.1038/s41477-025-02085-8. Online ahead of print. Nat Plants. 2025. PMID: 40897810
-
Novel Insights into the Landscape of Crossover and Noncrossover Events in Rhesus Macaques (Macaca mulatta).Genome Biol Evol. 2024 Jan 5;16(1):evad223. doi: 10.1093/gbe/evad223. Genome Biol Evol. 2024. PMID: 38051960 Free PMC article.
References
-
- Arnheim N, Calabrese P, Tiemann-Boege I. Mammalian meiotic recombination hot spots. Annu Rev Genet. 2007;41:369–399. - PubMed
-
- Baudat F, de Massy B. Regulating double-stranded DNA break repair towards crossover or non-crossover during mammalian meiosis. Chromosome Res. 2007b;15:565–577. - PubMed
-
- Baudat F, de Massy B. Parallel detection of crossovers and noncrossovers in mouse germ cells. Methods Mol Biol. 2009;557:305–322. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
