

Role of a ribosome-associated E3 ubiquitin ligase in protein quality control
- PMID: 20835226
- PMCID: PMC2988496
- DOI: 10.1038/nature09371
Role of a ribosome-associated E3 ubiquitin ligase in protein quality control
Abstract
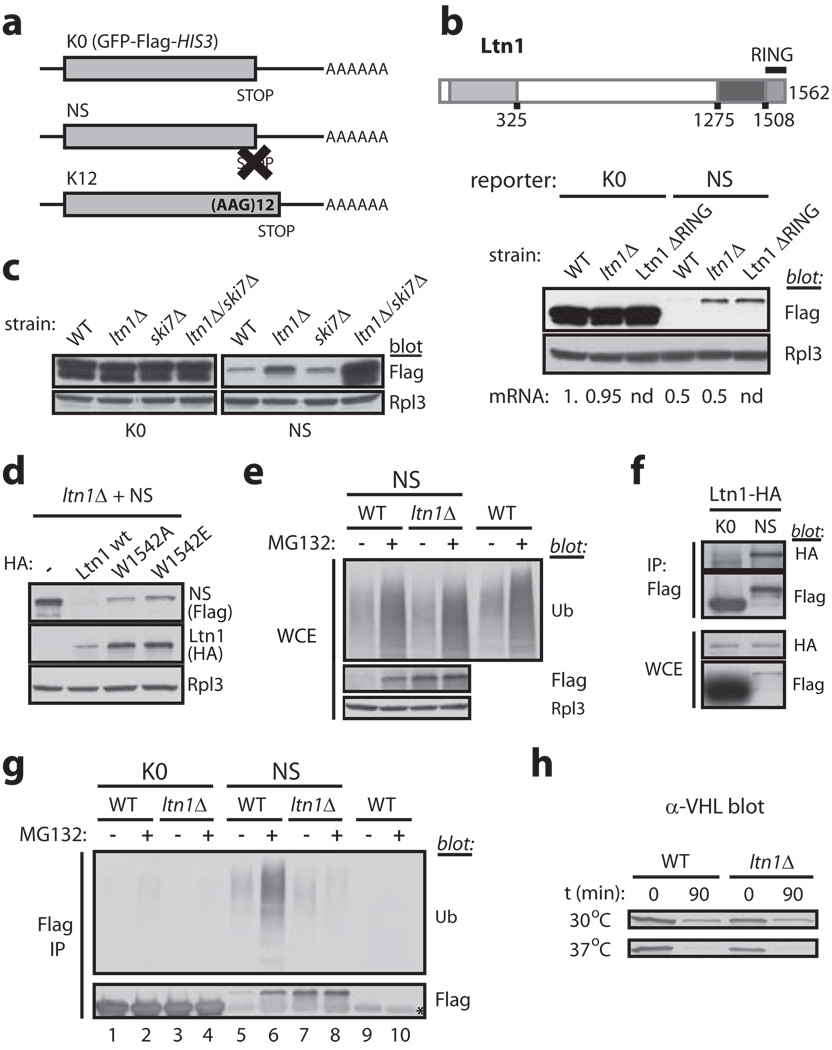
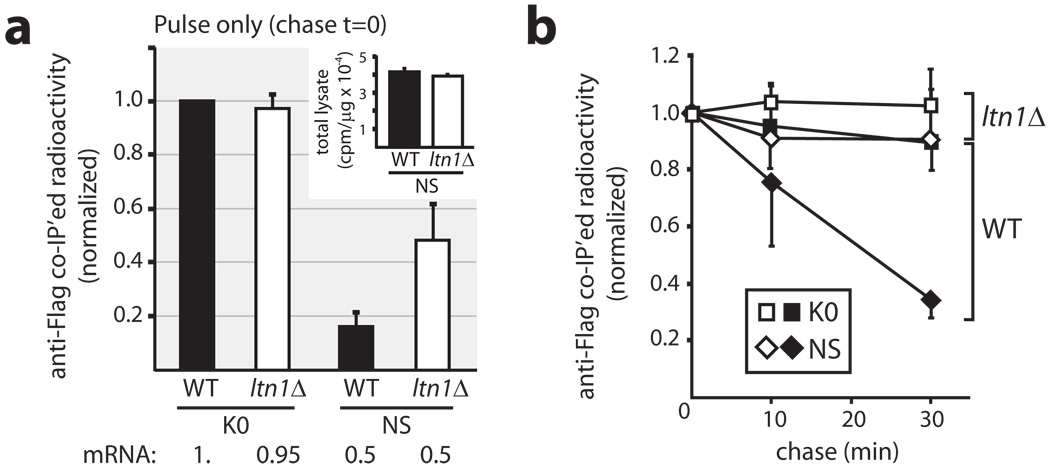
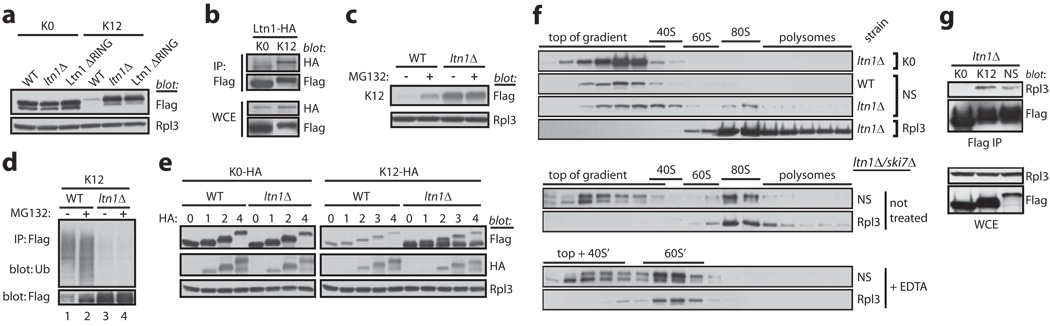
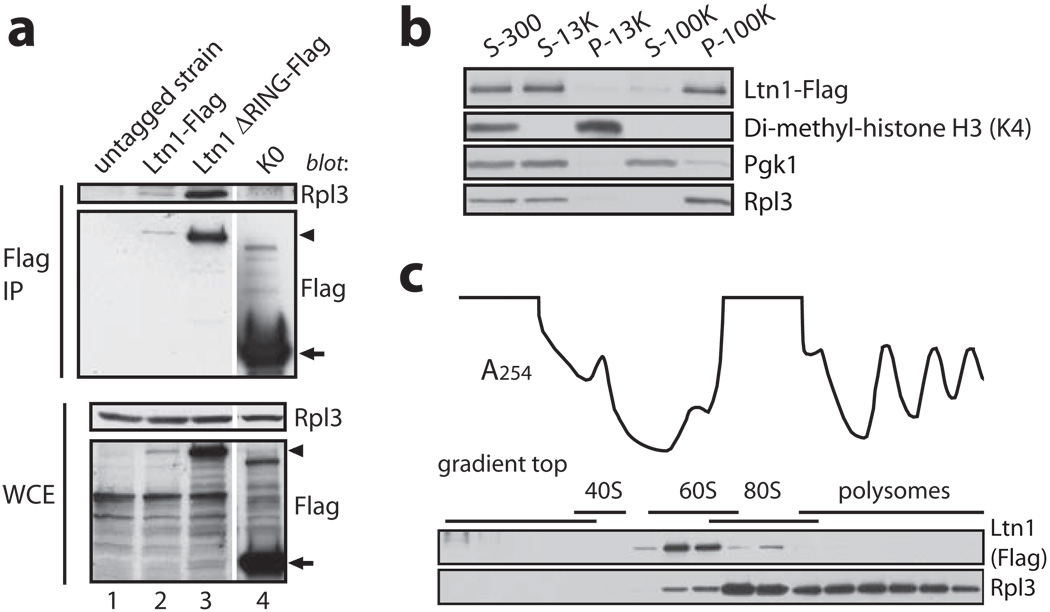
Messenger RNA lacking stop codons ('non-stop mRNA') can arise from errors in gene expression, and encode aberrant proteins whose accumulation could be deleterious to cellular function. In bacteria, these 'non-stop proteins' become co-translationally tagged with a peptide encoded by ssrA/tmRNA (transfer-messenger RNA), which signals their degradation by energy-dependent proteases. How eukaryotic cells eliminate non-stop proteins has remained unknown. Here we show that the Saccharomyces cerevisiae Ltn1 RING-domain-type E3 ubiquitin ligase acts in the quality control of non-stop proteins, in a process that is mechanistically distinct but conceptually analogous to that performed by ssrA: Ltn1 is predominantly associated with ribosomes, and it marks nascent non-stop proteins with ubiquitin to signal their proteasomal degradation. Ltn1-mediated ubiquitylation of non-stop proteins seems to be triggered by their stalling in ribosomes on translation through the poly(A) tail. The biological relevance of this process is underscored by the finding that loss of Ltn1 function confers sensitivity to stress caused by increased non-stop protein production. We speculate that defective protein quality control may underlie the neurodegenerative phenotype that results from mutation of the mouse Ltn1 homologue Listerin.
Figures
References
-
- Moore SD, Sauer RT. The tmRNA system for translational surveillance and ribosome rescue. Annu. Rev. Biochem. 2007;76:101–124. - PubMed
-
- Frischmeyer PA, van Hoof A, O'Donnell K, et al. An mRNA surveillance mechanism that eliminates transcripts lacking termination codons. Science. 2002;295:2258–2261. - PubMed
-
- Dulebohn D, Choy J, Sundermeier T, et al. Trans-translation: the tmRNA-mediated surveillance mechanism for ribosome rescue, directed protein degradation, and nonstop mRNA decay. Biochemistry. 2007;46:4681–4693. - PubMed
-
- van Hoof A, Frischmeyer PA, Dietz HC, et al. Exosome-mediated recognition and degradation of mRNAs lacking a termination codon. Science. 2002;295:2262–2264. - PubMed
-
- Doma MK, Parker R. RNA quality control in eukaryotes. Cell. 2007;131:660–668. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
