

Candidate exome capture identifies mutation of SDCCAG8 as the cause of a retinal-renal ciliopathy
- PMID: 20835237
- PMCID: PMC2947620
- DOI: 10.1038/ng.662
Candidate exome capture identifies mutation of SDCCAG8 as the cause of a retinal-renal ciliopathy
Abstract
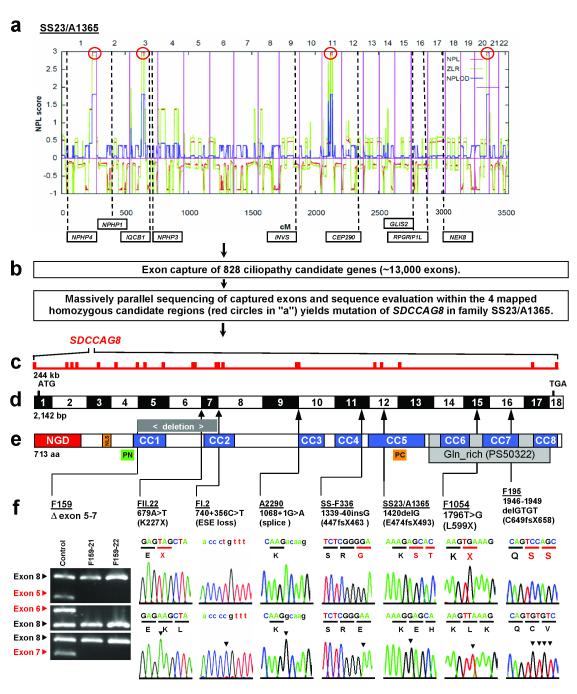
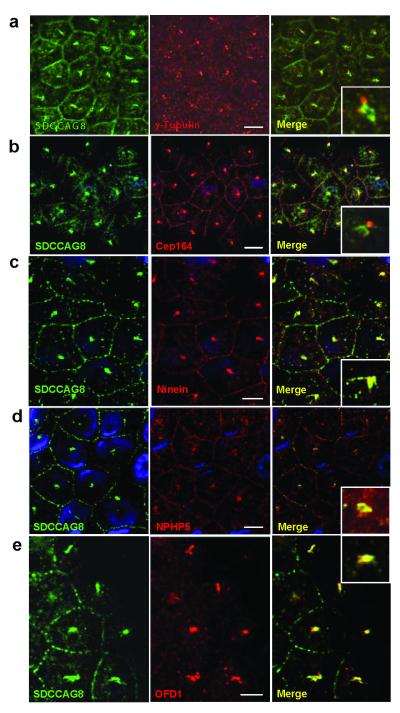
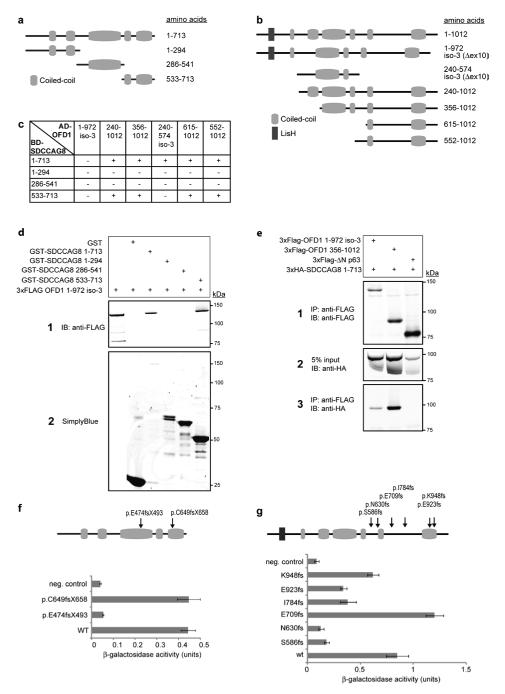
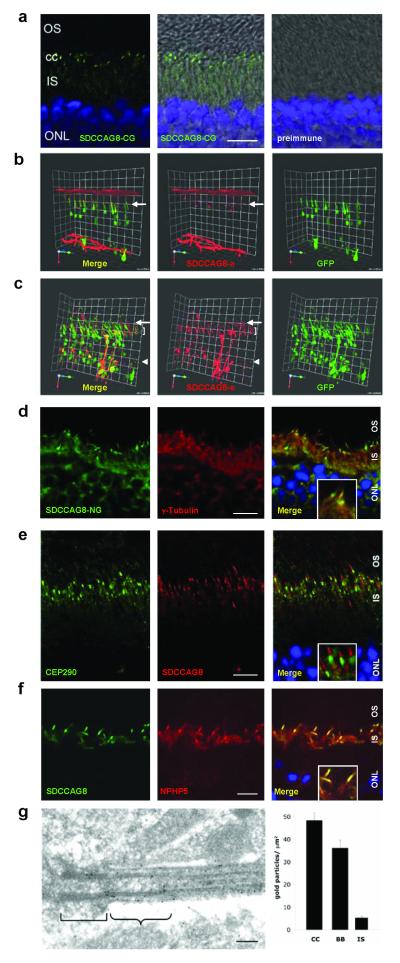
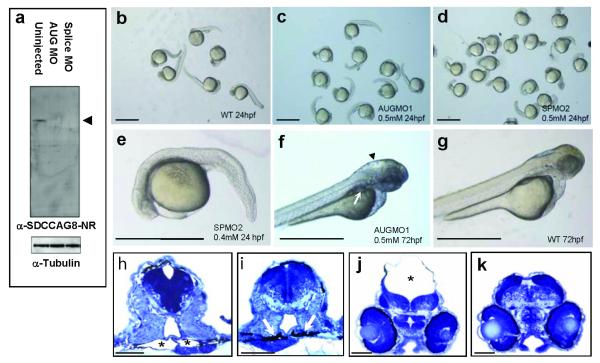
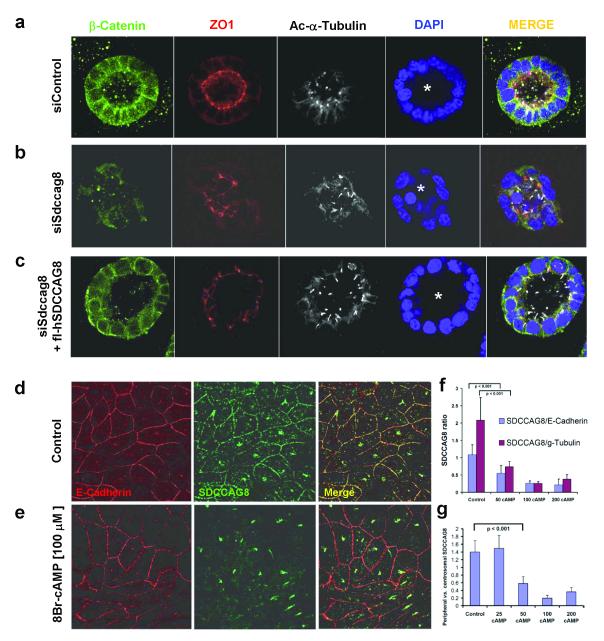
Nephronophthisis-related ciliopathies (NPHP-RC) are recessive disorders that feature dysplasia or degeneration occurring preferentially in the kidney, retina and cerebellum. Here we combined homozygosity mapping with candidate gene analysis by performing 'ciliopathy candidate exome capture' followed by massively parallel sequencing. We identified 12 different truncating mutations of SDCCAG8 (serologically defined colon cancer antigen 8, also known as CCCAP) in 10 families affected by NPHP-RC. We show that SDCCAG8 is localized at both centrioles and interacts directly with OFD1 (oral-facial-digital syndrome 1), which is associated with NPHP-RC. Depletion of sdccag8 causes kidney cysts and a body axis defect in zebrafish and induces cell polarity defects in three-dimensional renal cell cultures. This work identifies loss of SDCCAG8 function as a cause of a retinal-renal ciliopathy and validates exome capture analysis for broadly heterogeneous single-gene disorders.
Figures
Comment in
-
Mutations in a novel cilia-centrosome protein cause a cystic kidney disease associated with retinal degeneration.Clin Genet. 2011 Mar;79(3):222-4. doi: 10.1111/j.1399-0004.2010.01612.x. Epub 2011 Jan 4. Clin Genet. 2011. PMID: 21199495 No abstract available.
References
-
- Hildebrandt F, et al. A novel gene encoding an SH3 domain protein is mutated in nephronophthisis type 1. Nat Genet. 1997;17:149–153. - PubMed
-
- Olbrich H, et al. Mutations in a novel gene, NPHP3, cause adolescent nephronophthisis, tapeto-retinal degeneration and hepatic fibrosis. Nat Genet. 2003;34:455–459. - PubMed
-
- Mollet G, et al. The gene mutated in juvenile nephronophthisis type 4 encodes a novel protein that interacts with nephrocystin. Nat Genet. 2002;32:300–305. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
