

Non-native interactions play an effective role in protein folding dynamics
- PMID: 20836137
- PMCID: PMC3005790
- DOI: 10.1002/pro.498
Non-native interactions play an effective role in protein folding dynamics
Abstract
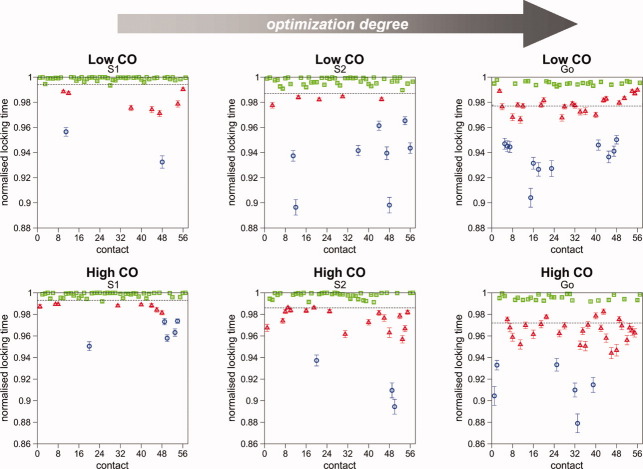
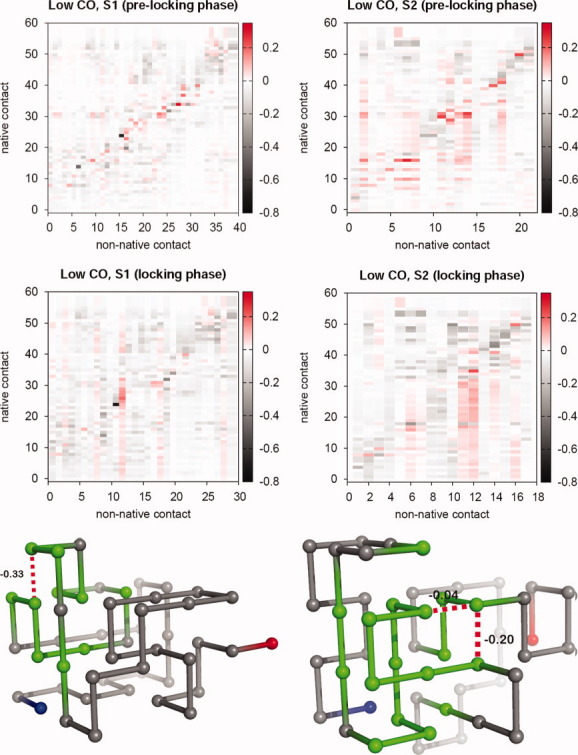
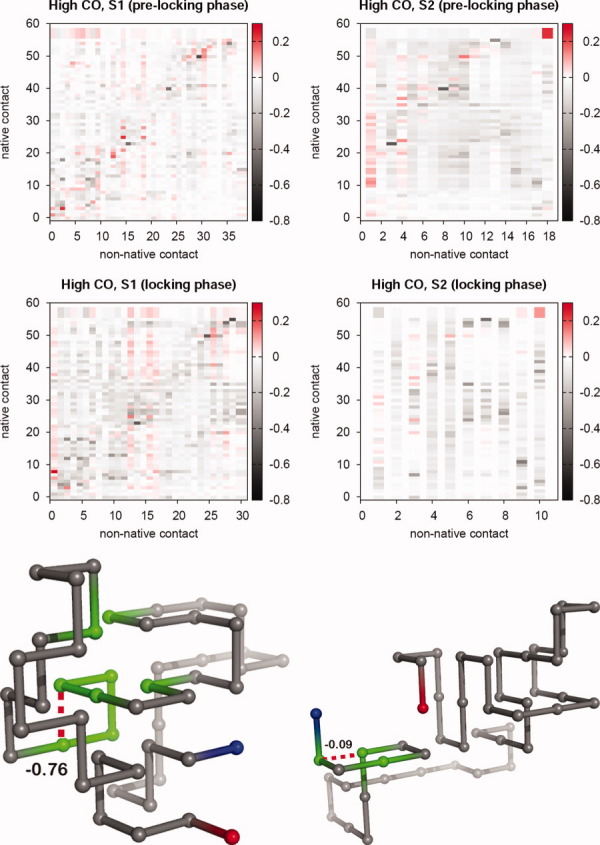
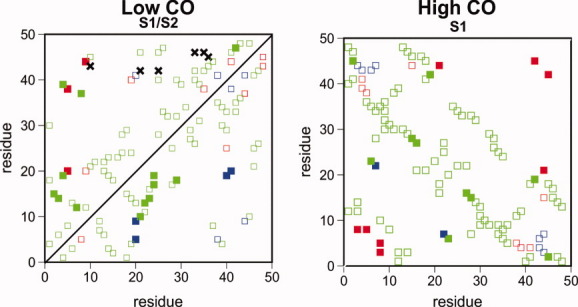
Systematic Monte Carlo simulations of simple lattice models show that the final stage of protein folding is an ordered process where native contacts get locked (i.e., the residues come into contact and remain in contact for the duration of the folding process) in a well-defined order. The detailed study of the folding dynamics of protein-like sequences designed as to exhibit different contact energy distributions, as well as different degrees of sequence optimization (i.e., participation of non-native interactions in the folding process), reveals significant differences in the corresponding locking scenarios--the collection of native contacts and their average locking times, which are largely ascribable to the dynamics of non-native contacts. Furthermore, strong evidence for a positive role played by non-native contacts at an early folding stage was also found. Interestingly, for topologically simple target structures, a positive interplay between native and non-native contacts is observed also toward the end of the folding process, suggesting that non-native contacts may indeed affect the overall folding process. For target models exhibiting clear two-state kinetics, the relation between the nucleation mechanism of folding and the locking scenario is investigated. Our results suggest that the stabilization of the folding transition state can be achieved through the establishment of a very small network of native contacts that are the first to lock during the folding process.
Figures
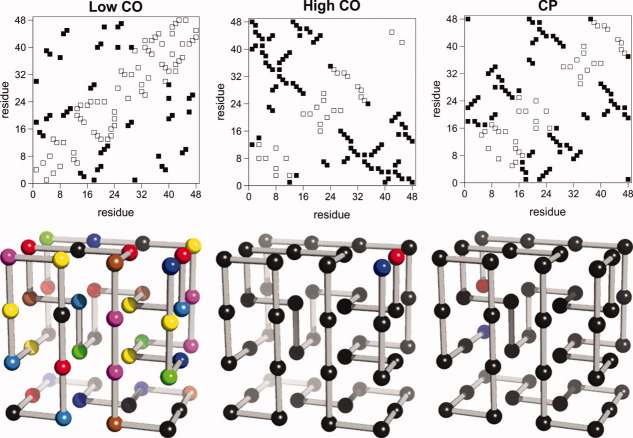
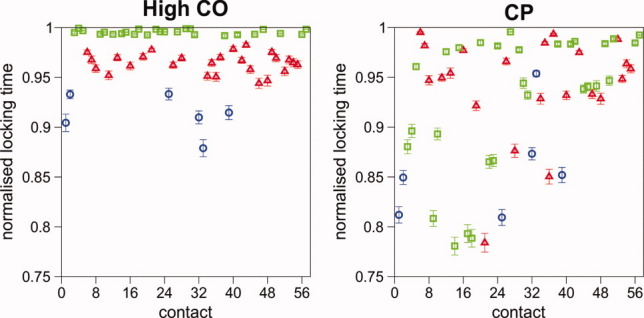
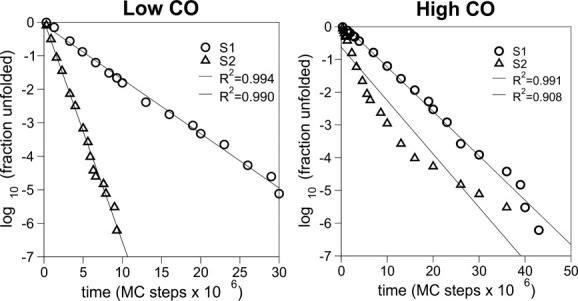
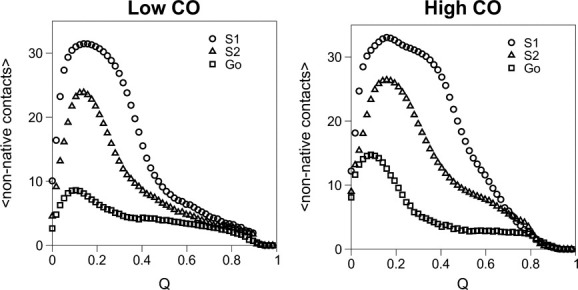
References
-
- Dill KA, Ozkan B, Weikl TR, Chodera JD, Voelz VA. The protein folding problem: when will it be solved? Curr Opin Struc Biol. 2007;17:342–346. - PubMed
-
- Service RF. Problem solved (*sort of) Science. 2008;321:784–786. - PubMed
-
- Jackson SE. How do small single-domain proteins fold? Fold Des. 1998;3:R81–R91. - PubMed
-
- Bai Y. Hidden intermediates and Levinthal paradox in the folding of small proteins. Biochem Biophys Res Commun. 2003;305:785–788. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
