

Role of matrix metalloproteinase-9 dimers in cell migration: design of inhibitory peptides
- PMID: 20837483
- PMCID: PMC2975217
- DOI: 10.1074/jbc.M109.091769
Role of matrix metalloproteinase-9 dimers in cell migration: design of inhibitory peptides
Abstract
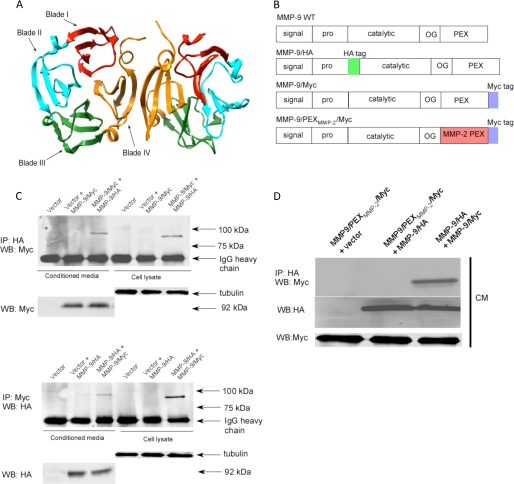
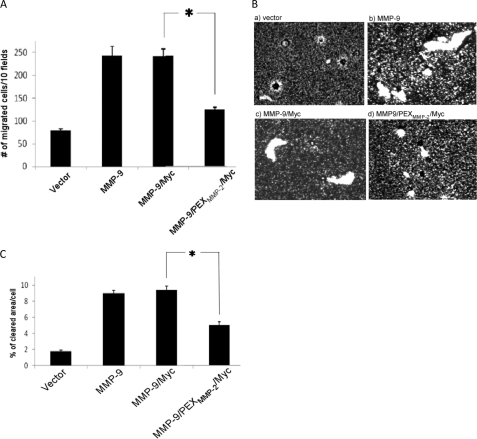
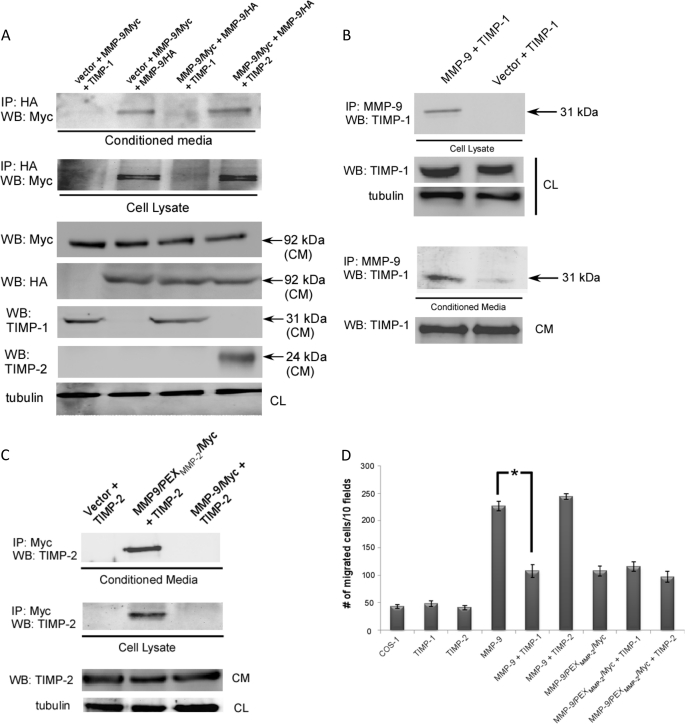
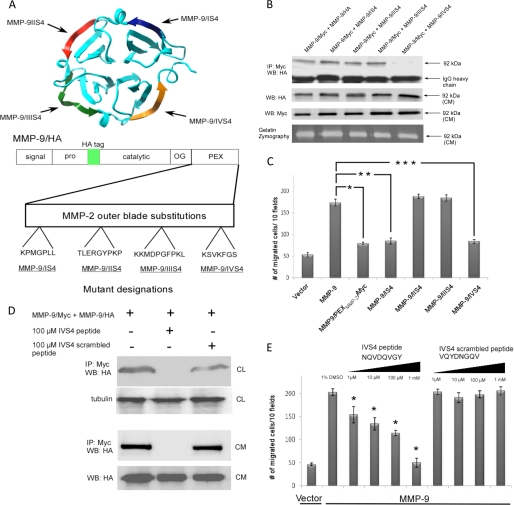
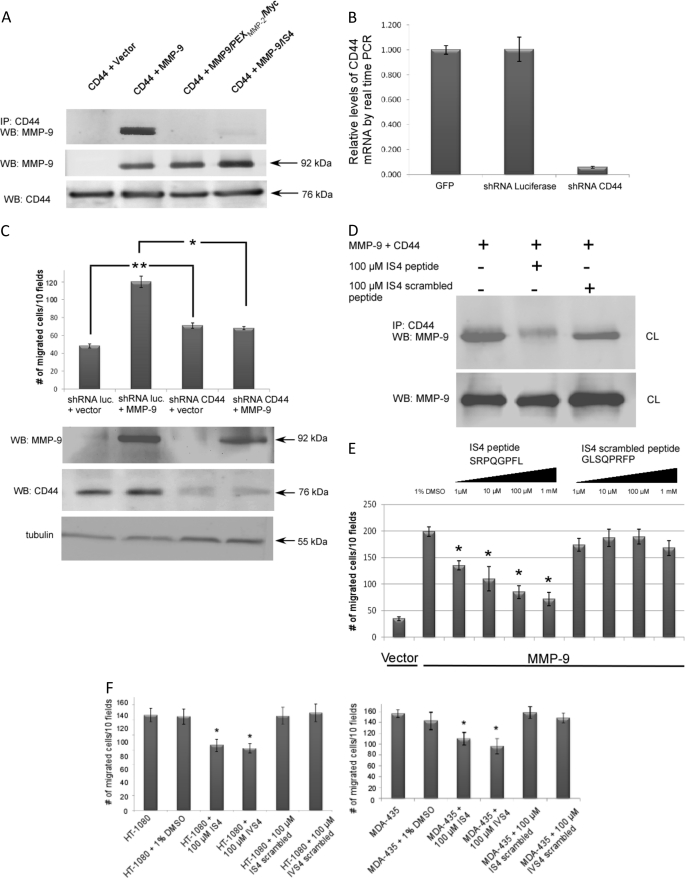
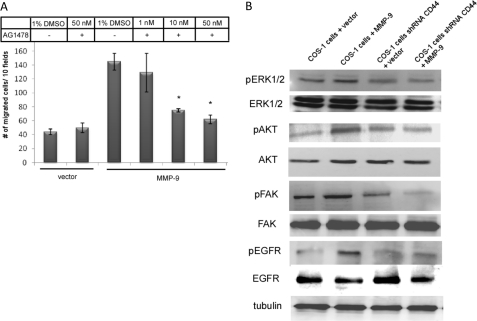
Non-proteolytic activities of matrix metalloproteinases (MMPs) have recently been shown to impact cell migration, but the precise mechanism remains to be understood. We previously demonstrated that the hemopexin (PEX) domain of MMP-9 is a prerequisite for enhanced cell migration. Using a biochemical approach, we now report that dimerization of MMP-9 through the PEX domain appears necessary for MMP-9-enhanced cell migration. Following a series of substitution mutations within the MMP-9 PEX domain, blade IV was shown to be critical for homodimerization, whereas blade I was required for heterodimerization with CD44. Blade I and IV mutants showed diminished enhancement of cell migration compared with wild type MMP-9-transfected cells. Peptides mimicking motifs in the outermost strands of the first and fourth blades of the MMP-9 PEX domain were designed. These peptides efficiently blocked MMP-9 dimer formation and inhibited motility of COS-1 cells overexpressing MMP-9, HT-1080, and MDA-MB-435 cells. Using a shRNA approach, CD44 was found to be a critical molecule in MMP-9-mediated cell migration. Furthermore, an axis involving a MMP-9-CD44-EGFR signaling pathway in cell migration was identified using antibody array and specific receptor tyrosine kinase inhibitors. In conclusion, we dissected the mechanism of pro-MMP-9-enhanced cell migration and developed structure-based inhibitory peptides targeting MMP-9-mediated cell migration.
Figures
Similar articles
-
Inhibition of matrix metalloproteinase 14 (MMP-14)-mediated cancer cell migration.J Biol Chem. 2011 Sep 23;286(38):33167-77. doi: 10.1074/jbc.M111.256644. Epub 2011 Jul 27. J Biol Chem. 2011. PMID: 21795678 Free PMC article.
-
Role of the hemopexin domain of matrix metalloproteinases in cell migration.J Cell Physiol. 2008 Dec;217(3):643-51. doi: 10.1002/jcp.21535. J Cell Physiol. 2008. PMID: 18636552 Free PMC article.
-
Small-molecule anticancer compounds selectively target the hemopexin domain of matrix metalloproteinase-9.Cancer Res. 2011 Jul 15;71(14):4977-88. doi: 10.1158/0008-5472.CAN-10-4552. Epub 2011 Jun 6. Cancer Res. 2011. PMID: 21646471 Free PMC article.
-
Electrochemical impedance spectroscopy for quantization of matrix Metalloproteinase-14 based on peptides inhibiting its homodimerization and heterodimerization.Talanta. 2019 Dec 1;205:120142. doi: 10.1016/j.talanta.2019.120142. Epub 2019 Jul 18. Talanta. 2019. PMID: 31450394 Review.
-
Biochemistry and molecular biology of gelatinase B or matrix metalloproteinase-9 (MMP-9): the next decade.Crit Rev Biochem Mol Biol. 2013 May-Jun;48(3):222-72. doi: 10.3109/10409238.2013.770819. Epub 2013 Apr 2. Crit Rev Biochem Mol Biol. 2013. PMID: 23547785 Review.
Cited by
-
A new isoform of steroid receptor coactivator-1 is crucial for pathogenic progression of endometriosis.Nat Med. 2012 Jul;18(7):1102-11. doi: 10.1038/nm.2826. Nat Med. 2012. PMID: 22660634 Free PMC article.
-
Gelatinase B/MMP-9 in Tumour Pathogenesis and Progression.Cancers (Basel). 2014 Jan 27;6(1):240-96. doi: 10.3390/cancers6010240. Cancers (Basel). 2014. PMID: 24473089 Free PMC article.
-
MMP-9-induced increase in intestinal epithelial tight permeability is mediated by p38 kinase signaling pathway activation of MLCK gene.Am J Physiol Gastrointest Liver Physiol. 2019 Feb 1;316(2):G278-G290. doi: 10.1152/ajpgi.00126.2018. Epub 2018 Dec 13. Am J Physiol Gastrointest Liver Physiol. 2019. PMID: 30543452 Free PMC article.
-
Role of L1 cell adhesion molecule (L1CAM) in the metastatic cascade: promotion of dissemination, colonization, and metastatic growth.Clin Exp Metastasis. 2014 Jan;31(1):87-100. doi: 10.1007/s10585-013-9613-6. Epub 2013 Sep 4. Clin Exp Metastasis. 2014. PMID: 24002299
-
Bifunctional Inhibitors as a New Tool To Reduce Cancer Cell Invasion by Impairing MMP-9 Homodimerization.ACS Med Chem Lett. 2017 Feb 7;8(3):293-298. doi: 10.1021/acsmedchemlett.6b00446. eCollection 2017 Mar 9. ACS Med Chem Lett. 2017. PMID: 28337319 Free PMC article.
References
-
- Morrison C. J., Butler G. S., Rodríguez D., Overall C. M. (2009) Curr. Opin. Cell. Biol. 21, 645–653 - PubMed
-
- Nagase H., Woessner J. F., Jr. (1999) J. Biol. Chem. 274, 21491–21494 - PubMed
-
- Björklund M., Koivunen E. (2005) Biochim. Biophys. Acta 1755, 37–69 - PubMed
-
- Zucker S., Cao J., Chen W. T. (2000) Oncogene 19, 6642–6650 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
