

Tafenoquine, an antiplasmodial 8-aminoquinoline, targets leishmania respiratory complex III and induces apoptosis
- PMID: 20837758
- PMCID: PMC2981264
- DOI: 10.1128/AAC.00790-10
Tafenoquine, an antiplasmodial 8-aminoquinoline, targets leishmania respiratory complex III and induces apoptosis
Abstract
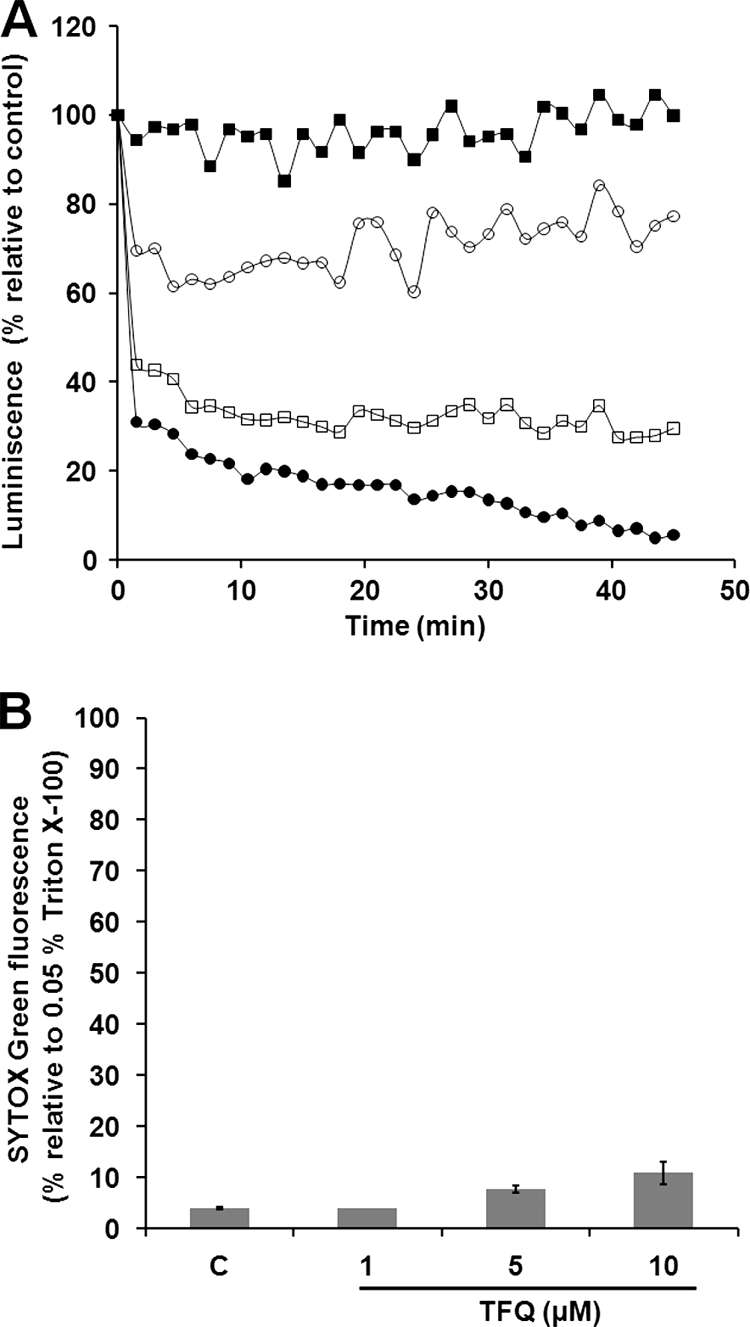
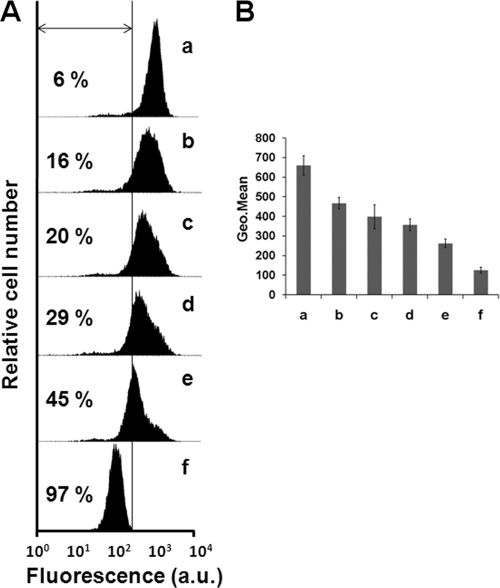
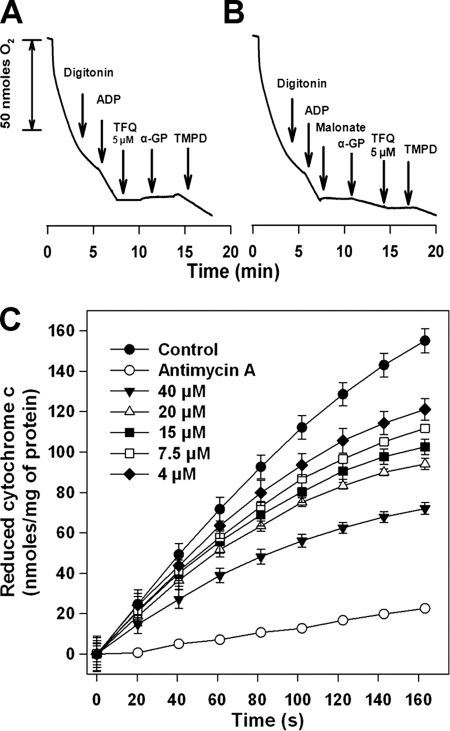
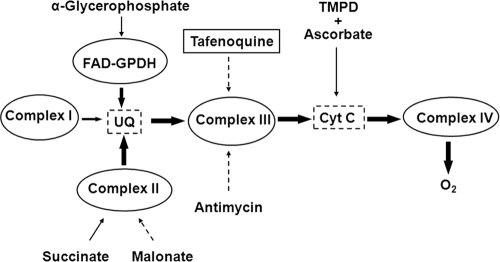
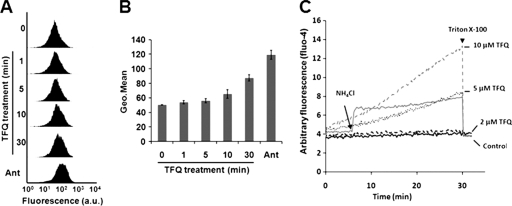
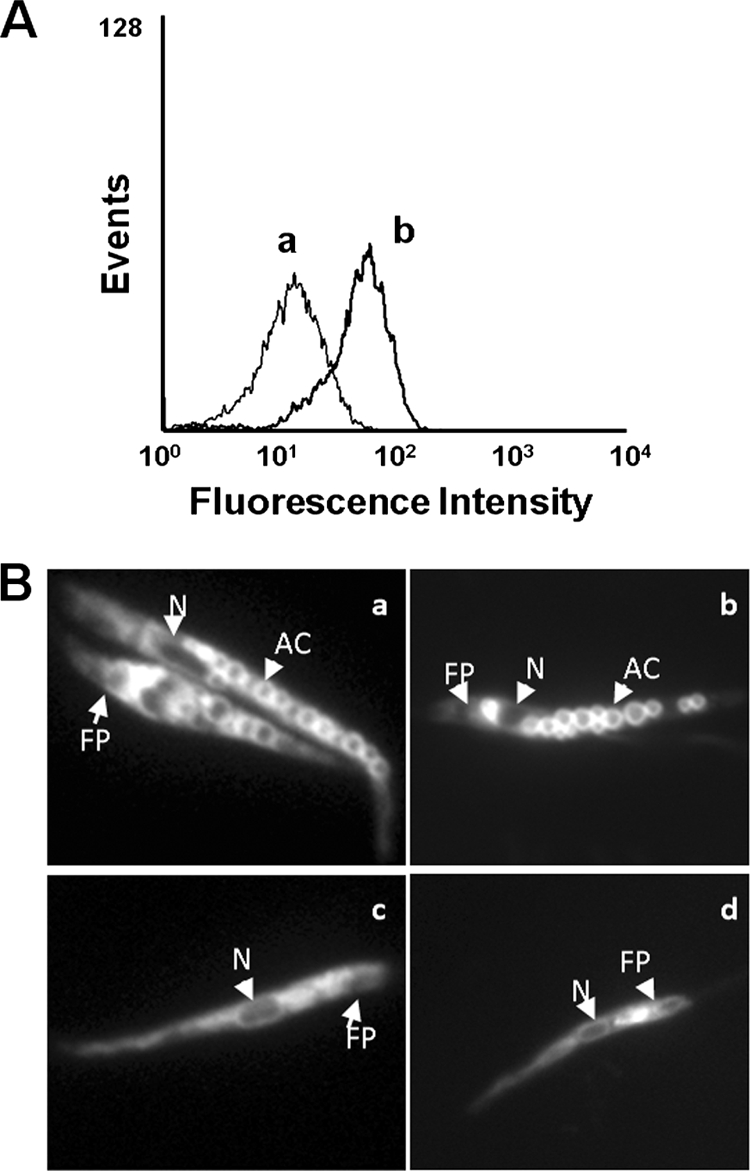
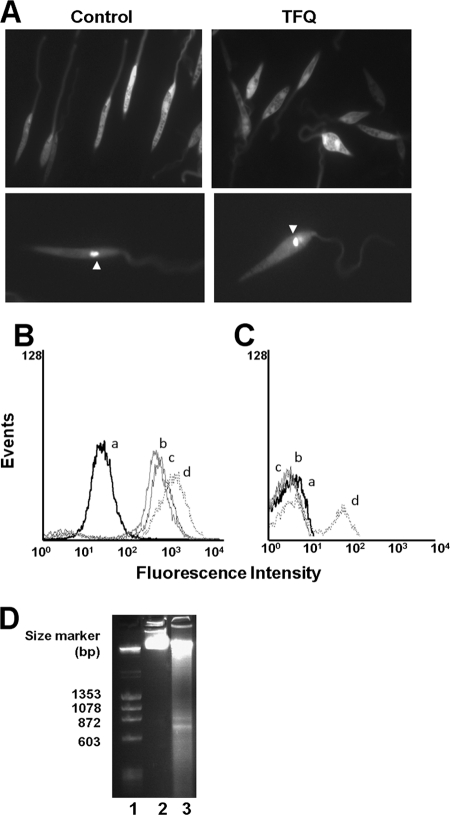
Tafenoquine (TFQ), an 8-aminoquinoline analogue of primaquine, which is currently under clinical trial (phase IIb/III) for the treatment and prevention of malaria, may represent an alternative treatment for leishmaniasis. In this work, we have studied the mechanism of action of TFQ against Leishmania parasites. TFQ impaired the overall bioenergetic metabolism of Leishmania promastigotes, causing a rapid drop in intracellular ATP levels without affecting plasma membrane permeability. TFQ induced mitochondrial dysfunction through the inhibition of cytochrome c reductase (respiratory complex III) with a decrease in the oxygen consumption rate and depolarization of mitochondrial membrane potential. This was accompanied by ROS production, elevation of intracellular Ca(2+) levels and concomitant nuclear DNA fragmentation. We conclude that TFQ targets Leishmania mitochondria, leading to an apoptosis-like death process.
Figures
References
-
- Alvarez-Fortes, E., L. M. Ruiz-Perez, F. Bouillaud, E. Rial, and L. Rivas. 1998. Expression and regulation of mitochondrial uncoupling protein 1 from brown adipose tissue in Leishmania major promastigotes. Mol. Biochem. Parasitol. 93:191-202. - PubMed
-
- Besteiro, S., D. Tonn, L. Tetley, G. H. Coombs, and J. C. Mottram. 2008. The AP3 adaptor is involved in the transport of membrane proteins to acidocalcisomes of Leishmania. J. Cell Sci. 121:561-570. - PubMed
-
- Besteiro, S., R. A. Williams, L. S. Morrison, G. H. Coombs, and J. C. Mottram. 2006. Endosome sorting and autophagy are essential for differentiation and virulence of Leishmania major. J. Biol. Chem. 281:11384-11396. - PubMed
-
- Chappuis, F., S. Sundar, A. Hailu, H. Ghalib, S. Rijal, R. W. Peeling, J. Alvar, and M. Boelaert. 2007. Visceral leishmaniasis: what are the needs for diagnosis, treatment and control? Nat. Rev. Microbiol. 5:873-882. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
