

Small-molecule antioxidant proteome-shields in Deinococcus radiodurans
- PMID: 20838443
- PMCID: PMC2933237
- DOI: 10.1371/journal.pone.0012570
Small-molecule antioxidant proteome-shields in Deinococcus radiodurans
Abstract
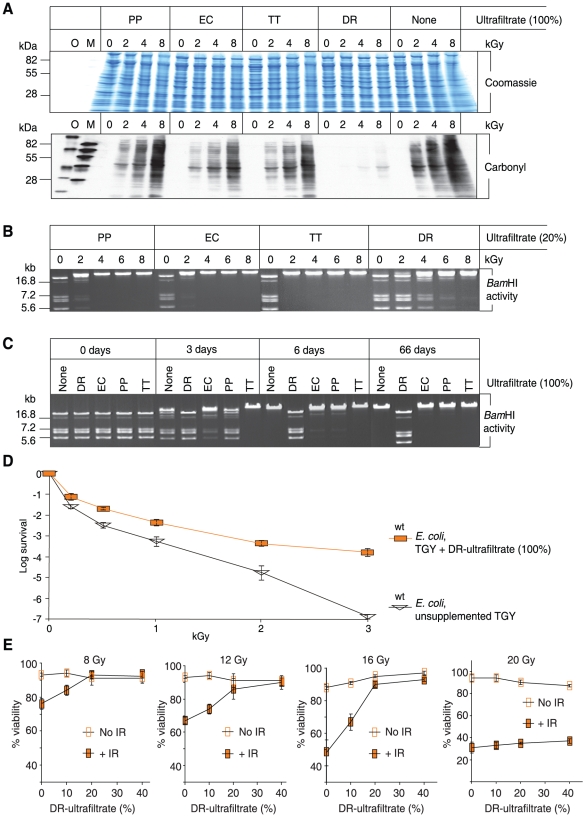
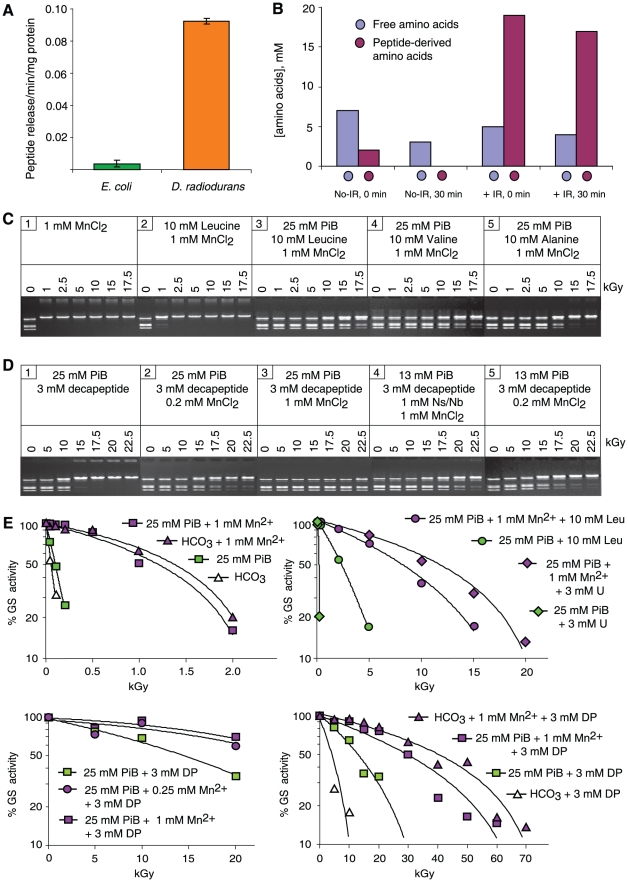
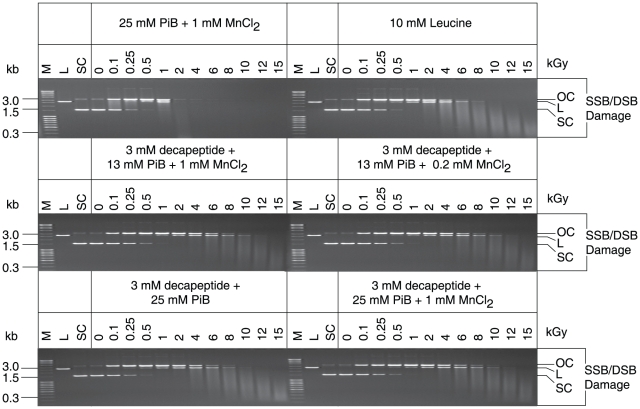
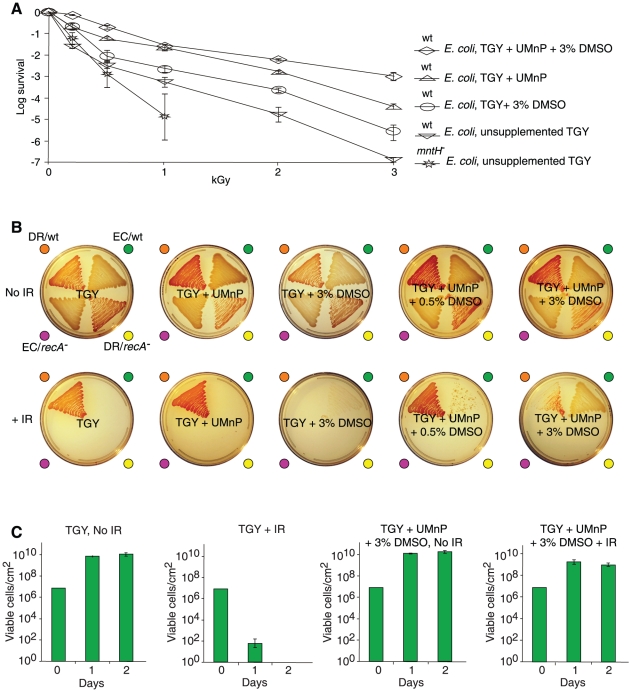
For Deinococcus radiodurans and other bacteria which are extremely resistant to ionizing radiation, ultraviolet radiation, and desiccation, a mechanistic link exists between resistance, manganese accumulation, and protein protection. We show that ultrafiltered, protein-free preparations of D. radiodurans cell extracts prevent protein oxidation at massive doses of ionizing radiation. In contrast, ultrafiltrates from ionizing radiation-sensitive bacteria were not protective. The D. radiodurans ultrafiltrate was enriched in Mn, phosphate, nucleosides and bases, and peptides. When reconstituted in vitro at concentrations approximating those in the D. radiodurans cytosol, peptides interacted synergistically with Mn(2+) and orthophosphate, and preserved the activity of large, multimeric enzymes exposed to 50,000 Gy, conditions which obliterated DNA. When applied ex vivo, the D. radiodurans ultrafiltrate protected Escherichia coli cells and human Jurkat T cells from extreme cellular insults caused by ionizing radiation. By establishing that Mn(2+)-metabolite complexes of D. radiodurans specifically protect proteins against indirect damage caused by gamma-rays delivered in vast doses, our findings provide the basis for a new approach to radioprotection and insight into how surplus Mn budgets in cells combat reactive oxygen species.
Conflict of interest statement
Figures


References
-
- Kriško A, Radman M. Protein damage and death by radiation in Escherichia coli and Deinococcus radiodurans. Proc Natl Acad Sci USA. 2010 ( www.pnas.org/cgi/doi/10.1073/pnas.1009312107) - DOI - PMC - PubMed
-
- Daly MJ, Gaidamakova EK, Matrosova VY, Vasilenko A, Zhai M, et al. Accumulation of Mn(II) in Deinococcus radiodurans facilitates gamma-radiation resistance. Science. 2004;306:1025–1028. - PubMed
-
- Daly MJ. A new perspective on radiation resistance based on Deinococcus radiodurans. Nat Rev Microbiol. 2009;7:237–245. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
