

The role of calcium release activated calcium channels in osteoclast differentiation
- PMID: 20839232
- PMCID: PMC4020518
- DOI: 10.1002/jcp.22423
The role of calcium release activated calcium channels in osteoclast differentiation
Abstract
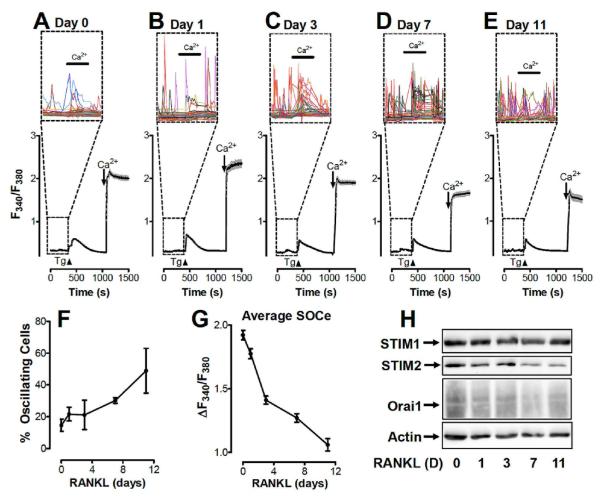
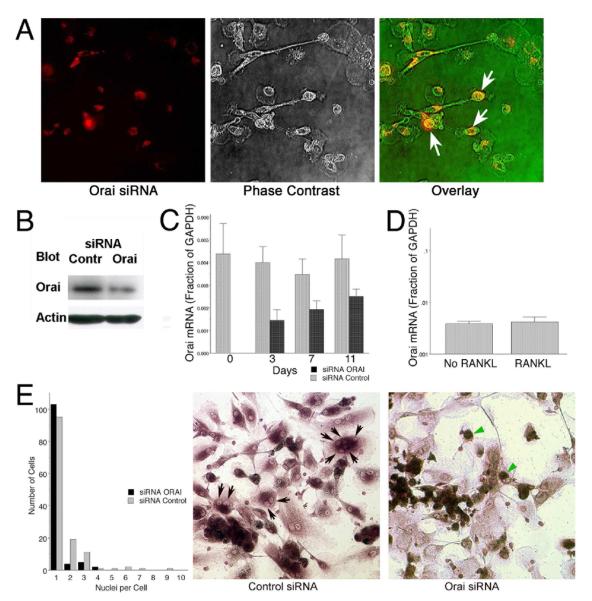
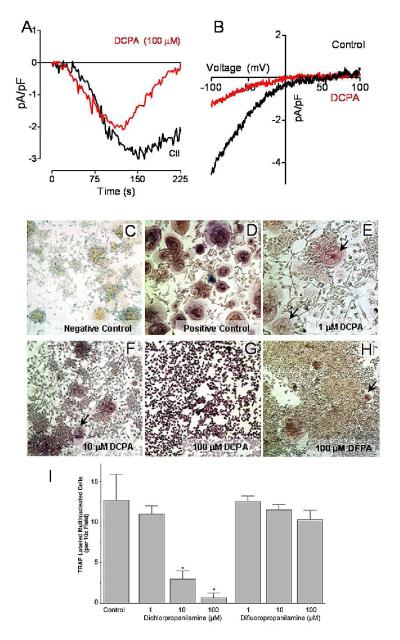
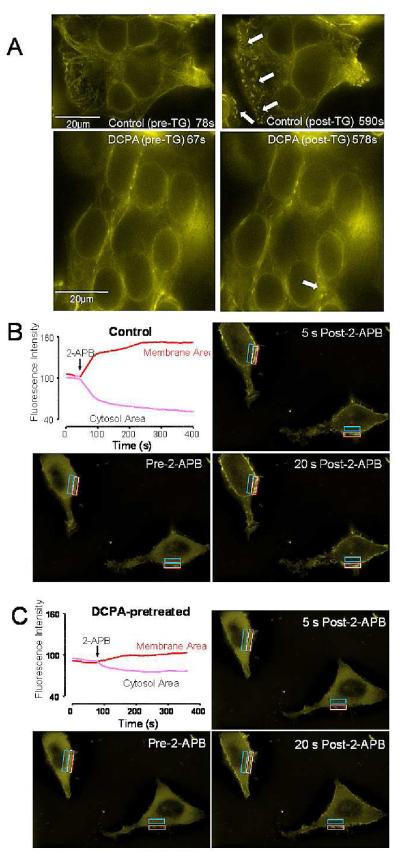
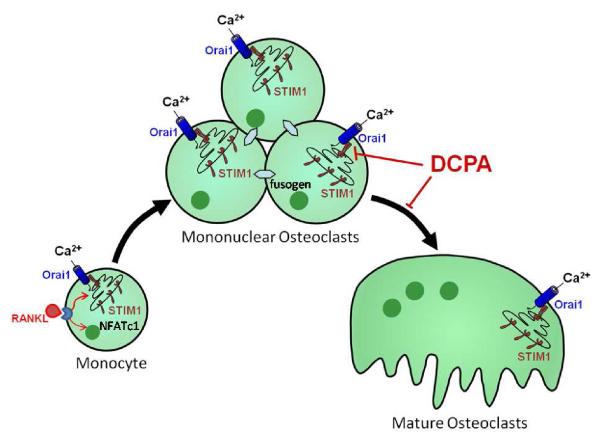
Osteoclasts are specialized macrophage derivatives that secrete acid and proteinases to mobilize bone for mineral homeostasis, growth, and replacement or repair. Osteoclast differentiation generally requires the monocyte growth factor m-CSF and the TNF-family cytokine RANKL, although differentiation is regulated by many other cytokines and by intracellular signals, including Ca(2+). Studies of osteoclast differentiation in vitro were performed using human monocytic precursors stimulated with m-CSF and RANKL, revealing significant loss in both the expression and function of the required components of store-operated Ca(2+) entry over the course of osteoclast differentiation. However, inhibition of CRAC using either the pharmacological agent 3,4-dichloropropioanilide (DCPA) or by knockdown of Orai1 severely inhibited formation of multinucleated osteoclasts. In contrast, no effect of CRAC channel inhibition was observed on expression of the osteoclast protein tartrate resistant acid phosphatase (TRAP). Our findings suggest that despite the fact that they are down-regulated during osteoclast differentiation, CRAC channels are required for cell fusion, a late event in osteoclast differentiation. Since osteoclasts cannot function properly without multinucleation, selective CRAC inhibitors may have utility in management of hyperresorptive states.
Copyright © 2010 Wiley-Liss, Inc.
Figures
References
-
- Chang EJ, Ha J, Huang H, Kim HJ, Woo JH, Lee Y, Lee ZH, Kim JH, Kim HH. The JNK-dependent CaMK pathway restrains the reversion of committed cells during osteoclast differentiation. Journal of Cell Science. 2008;121:2555–2564. - PubMed
-
- Day CJ, Kim MS, Lopez CM, Nicholson GC, Morrison NA. NFAT expression in human osteoclasts. J Cell Biochem. 2005;95:17–23. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
