

Evidence for the slow reaction of hypoxia-inducible factor prolyl hydroxylase 2 with oxygen
- PMID: 20840591
- PMCID: PMC4160827
- DOI: 10.1111/j.1742-4658.2010.07804.x
Evidence for the slow reaction of hypoxia-inducible factor prolyl hydroxylase 2 with oxygen
Abstract
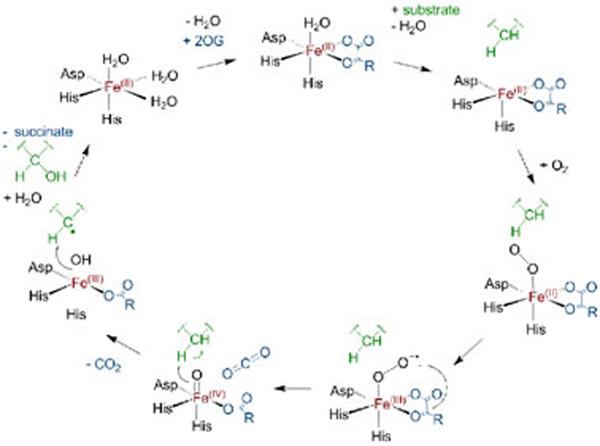
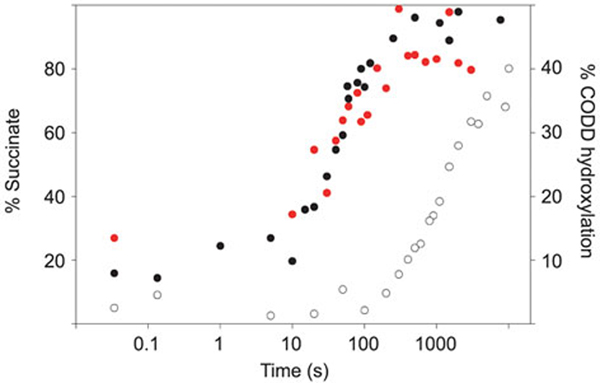
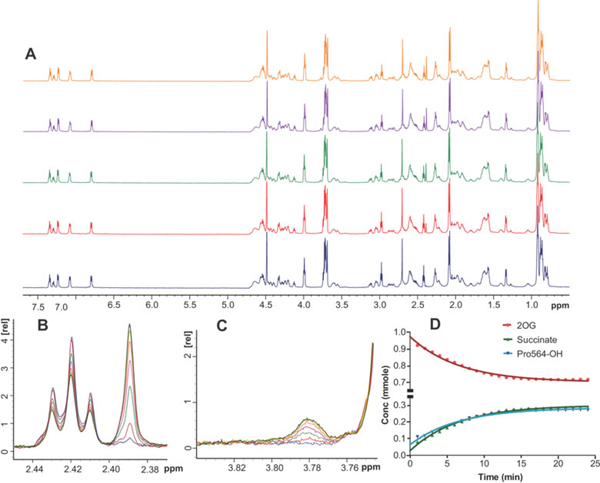
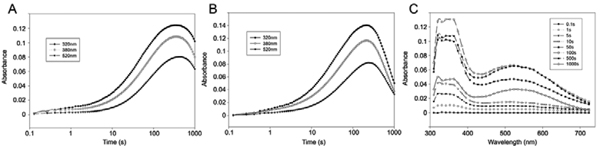
The response of animals to hypoxia is mediated by the hypoxia-inducible transcription factor. Human hypoxia-inducible factor is regulated by four Fe(II)- and 2-oxoglutarate-dependent oxygenases: prolyl hydroxylase domain enzymes 1-3 catalyse hydroxylation of two prolyl-residues in hypoxia-inducible factor, triggering its degradation by the proteasome. Factor inhibiting hypoxia-inducible factor catalyses the hydroxylation of an asparagine-residue in hypoxia-inducible factor, inhibiting its transcriptional activity. Collectively, the hypoxia-inducible factor hydroxylases negatively regulate hypoxia-inducible factor in response to increasing oxygen concentration. Prolyl hydroxylase domain 2 is the most important oxygen sensor in human cells; however, the underlying kinetic basis of the oxygen-sensing function of prolyl hydroxylase domain 2 is unclear. We report analyses of the reaction of prolyl hydroxylase domain 2 with oxygen. Chemical quench/MS experiments demonstrate that reaction of a complex of prolyl hydroxylase domain 2, Fe(II), 2-oxoglutarate and the C-terminal oxygen-dependent degradation domain of hypoxia-inducible factor-α with oxygen to form hydroxylated C-terminal oxygen-dependent degradation domain and succinate is much slower (approximately 100-fold) than for other similarly studied 2-oxoglutarate oxygenases. Stopped flow/UV-visible spectroscopy experiments demonstrate that the reaction produces a relatively stable species absorbing at 320 nm; Mössbauer spectroscopic experiments indicate that this species is likely not a Fe(IV)=O intermediate, as observed for other 2-oxoglutarate oxygenases. Overall, the results obtained suggest that, at least compared to other studied 2-oxoglutarate oxygenases, prolyl hydroxylase domain 2 reacts relatively slowly with oxygen, a property that may be associated with its function as an oxygen sensor.
© 2010 The Authors Journal compilation © 2010 FEBS.
Conflict of interest statement
Conflict of Interest: Prof. C.J. Schofield is a co-founder of ReOx Ltd., a company working on the exploitation of the hypoxic response.
Figures
References
-
- Kaelin WG, Jr, Ratcliffe PJ. Oxygen sensing by metazoans: the central role of the HIF hydroxylase pathway. Mol Cell. 2008;30:393–402. - PubMed
-
- Schofield CJ, Ratcliffe PJ. Oxygen sensing by HIF hydroxylases. Nat Rev Mol Cell Biol. 2004;5:343–354. - PubMed
-
- Semenza GL. Hydroxylation of HIF-1: oxygen sensing at the molecular level. Physiology (Bethesda) 2004;19:176–182. - PubMed
-
- Wang GL, Semenza GL. Purification and characterization of hypoxia-inducible factor-1. J Biol Chem. 1995;270:1230–1237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
