

Neutral antibodies to the TSH receptor are present in Graves' disease and regulate selective signaling cascades
- PMID: 20844004
- PMCID: PMC2954721
- DOI: 10.1210/en.2010-0424
Neutral antibodies to the TSH receptor are present in Graves' disease and regulate selective signaling cascades
Abstract
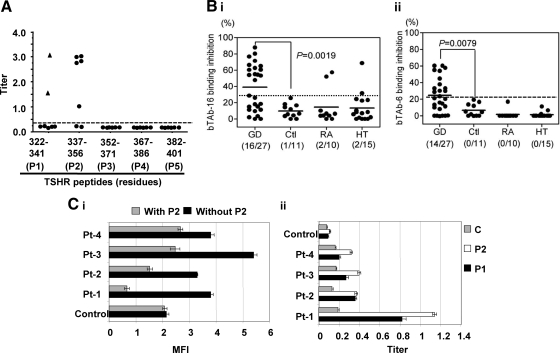
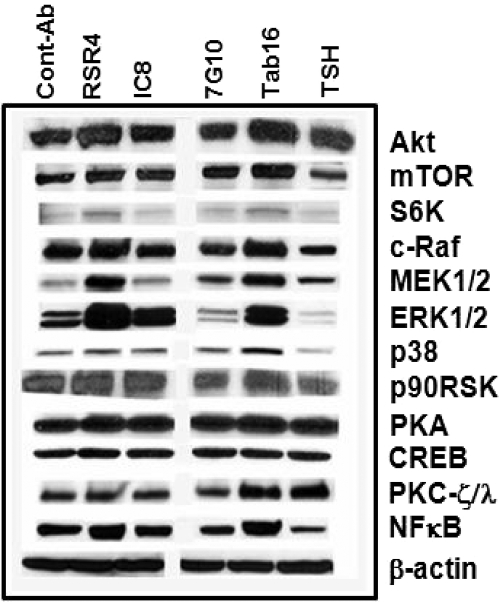
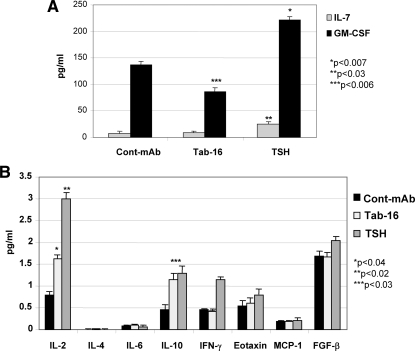
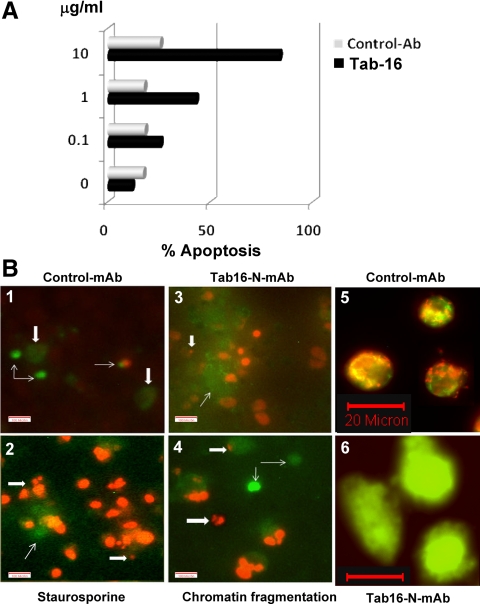
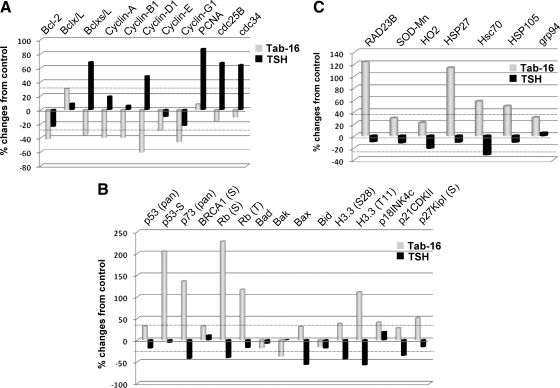
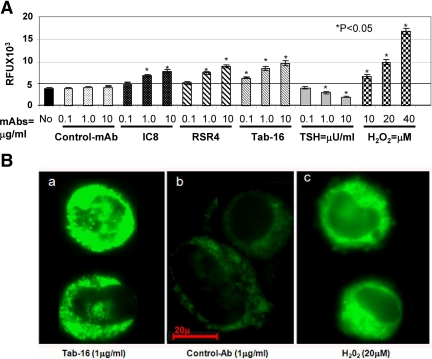
TSH receptor (TSHR) antibodies (Abs) may be stimulating, blocking, or neutral in their functional influences and are found in patients with autoimmune thyroid disease, especially Graves' disease (GD). Stimulators are known to activate the thyroid epithelial cells via both Gs- and Gq-coupled signaling pathways, whereas blockers inhibit the action of TSH and may act as weak agonists. However, TSHR neutral Abs do not block TSH binding and are unable to induce cAMP via Gsα. The importance of such neutral Abs in GD remains unclear because their functional consequence has been assumed to be zero. We hypothesized that: 1) neutral TSHR Abs are more common to GD than generally recognized; 2) they may induce distinct signaling imprints at the TSHR not seen with TSH itself; and 3) these signaling events may alter cellular function. To evaluate these hypotheses, we first confirmed the presence of neutral TSHR Abs in sera from patients with GD and then, using mouse and hamster neutral TSHR monoclonal Abs (N-mAbs) performed detailed signaling studies, including a proteomic Ab array, with rat thyrocytes (FRTL-5) as targets. This allowed us to examine a battery of signaling cascades and their downstream effectors. Neutral TSHR Abs were indeed frequently present in sera from patients with GD. Sixteen of 27 patients (59%) had detectable neutral TSHR Abs by competition assay with N-mAbs. On examining signaling cascades, we found that N-mAbs induced signal transduction, primarily via the protein kinase A II cascade. In addition to the activation of phosphatidylinositol 3K/Akt, N-mAbs, unlike TSH, had the ability to exclusively activate the mammalian target of rapamycin/p70 S6K, nuclear factor-κB, and MAPK-ERK1/2/p38α signaling cascades and their downstream effectors p90 ribosomal kinase/MAPK-interacting kinase-1/mitogen and stress-activated kinase-1 and N-mAbs activated all forms of protein kinase C isozymes. To define the downstream effector mechanisms produced by these signaling cascades, cytokine production, proliferation, and apoptosis in thyrocytes were investigated. Although N-mAbs produced less cytokines and proliferation compared with TSH, they had the distinction of inducing thyroid cell apoptosis under the experimental conditions used. When dissecting out possible mechanisms of apoptosis, we found that activation of multiple oxidative stress markers was the primary mechanism orchestrating the death signals. Therefore, using oxidative stress-induced apoptosis, N-mAbs may be capable of exacerbating the autoimmune response in GD via apoptotic cells inducing antigen-driven mechanisms. This may help explain the inflammatory nature of this common disorder.
Figures
References
-
- Kimura T, Van Keymeulen A, Golstein J, Fusco A, Dumont JE, Roger PP 2001 Regulation of thyroid cell proliferation by TSH and other factors: a critical evaluation of in vitro models. Endocr Rev 22:631–656 - PubMed
-
- Medina DL, Santisteban P 2000 Thyrotropin-dependent proliferation of in vitro rat thyroid cell systems. Eur J Endocrinol 143:161–178 - PubMed
-
- Ando T, Latif R, Daniel S, Eguchi K, Davies TF 2004 Dissecting linear and conformational epitopes on the native thyrotropin receptor. Endocrinology 145:5185–5193 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
