

Centriolar association of ALMS1 and likely centrosomal functions of the ALMS motif-containing proteins C10orf90 and KIAA1731
- PMID: 20844083
- PMCID: PMC2965680
- DOI: 10.1091/mbc.E10-03-0246
Centriolar association of ALMS1 and likely centrosomal functions of the ALMS motif-containing proteins C10orf90 and KIAA1731
Abstract
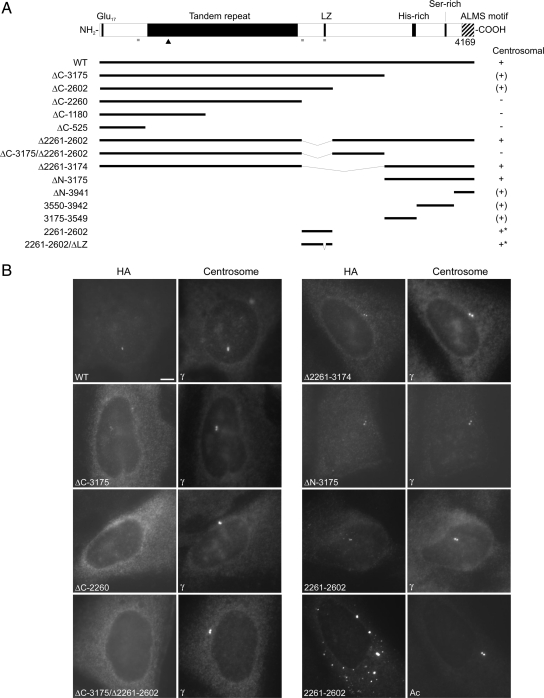
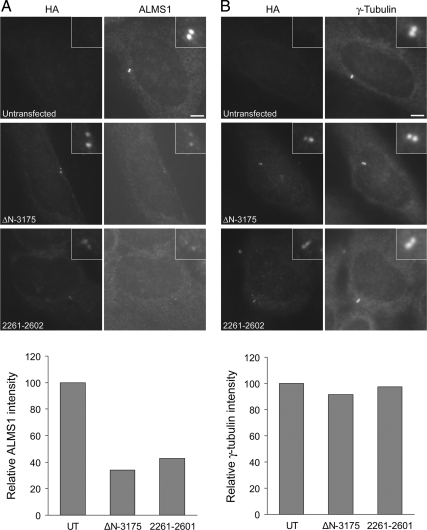
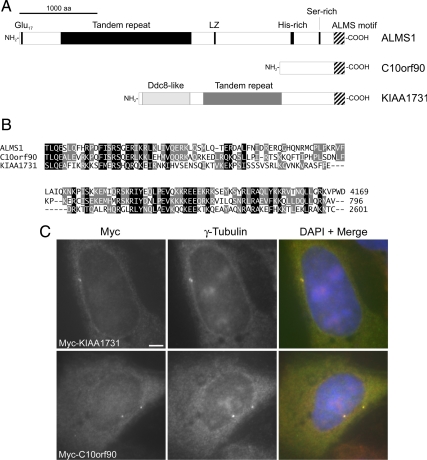
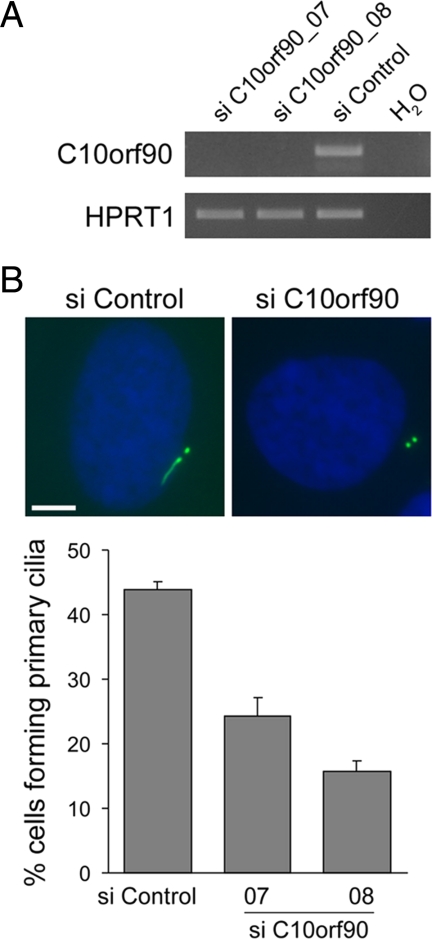
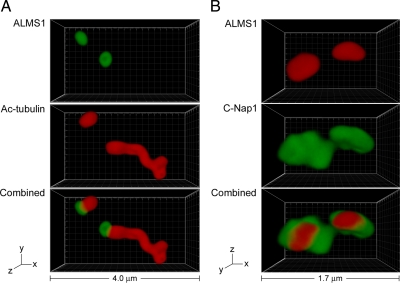
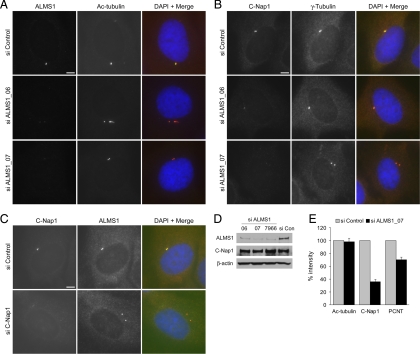
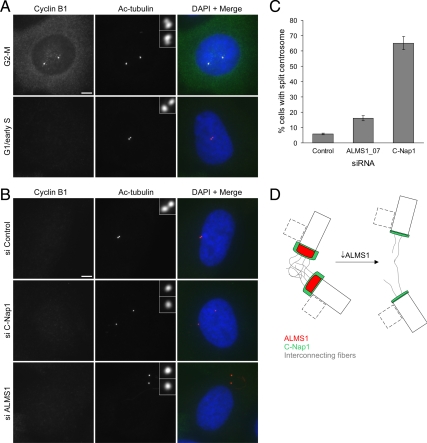
Mutations in the human gene ALMS1 cause Alström syndrome, a rare progressive condition characterized by neurosensory degeneration and metabolic defects. ALMS1 protein localizes to the centrosome and has been implicated in the assembly and/or maintenance of primary cilia; however its precise function, distribution within the centrosome, and mechanism of centrosomal recruitment are unknown. The C-terminus of ALMS1 contains a region with similarity to the uncharacterized human protein C10orf90, termed the ALMS motif. Here, we show that a third human protein, the candidate centrosomal protein KIAA1731, contains an ALMS motif and that exogenously expressed KIAA1731 and C10orf90 localize to the centrosome. However, based on deletion analysis of ALMS1, the ALMS motif appears unlikely to be critical for centrosomal targeting. RNAi analyses suggest that C10orf90 and KIAA1731 have roles in primary cilium assembly and centriole formation/stability, respectively. We also show that ALMS1 localizes specifically to the proximal ends of centrioles and basal bodies, where it colocalizes with the centrosome cohesion protein C-Nap1. RNAi analysis reveals markedly diminished centrosomal levels of C-Nap1 and compromised cohesion of parental centrioles in ALMS1-depleted cells. In summary, these data suggest centrosomal functions for C10orf90 and KIAA1731 and new centriole-related functions for ALMS1.
Figures


References
-
- Alstrom C. H., Hallgren B., Nilsson L. B., Asander H. Retinal degeneration combined with obesity, diabetes mellitus and neurogenous deafness: a specific syndrome (not hitherto described) distinct from the Laurence-Moon-Bardet-Biedl syndrome: a clinical, endocrinological and genetic examination based on a large pedigree. Acta Psychiatr. Neurol. Scand. 1959;34:1–35. - PubMed
-
- Andersen J. S., Wilkinson C. J., Mayor T., Mortensen P., Nigg E. A., Mann M. Proteomic characterization of the human centrosome by protein correlation profiling. Nature. 2003;426:570–574. - PubMed
-
- Azimzadeh J., Bornens M. Structure and duplication of the centrosome. J. Cell Sci. 2007;120:2139–2142. - PubMed
-
- Badano J. L., Mitsuma N., Beales P. L., Katsanis N. The ciliopathies: an emerging class of human genetic disorders. Annu. Rev. Genom. Hum. Genet. 2006;7:125–148. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
