

Pathogenic lysosomal depletion in Parkinson's disease
- PMID: 20844148
- PMCID: PMC6633458
- DOI: 10.1523/JNEUROSCI.1920-10.2010
Pathogenic lysosomal depletion in Parkinson's disease
Abstract
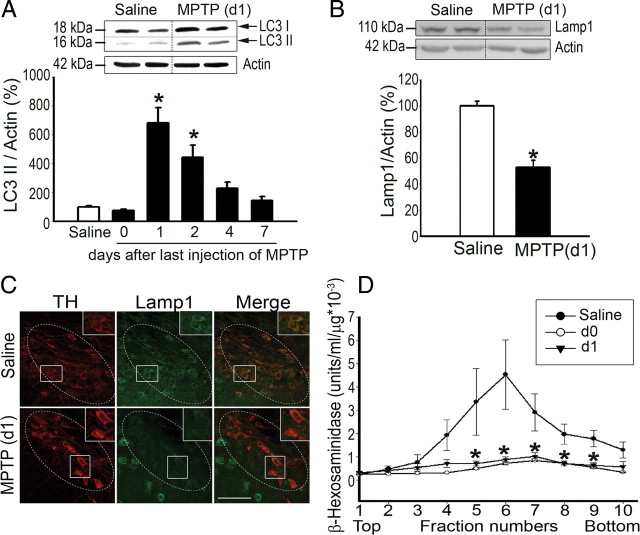
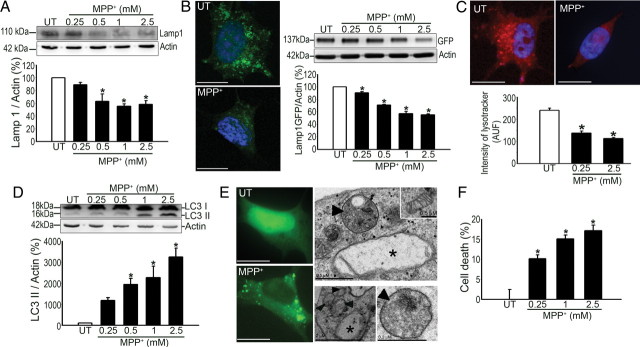
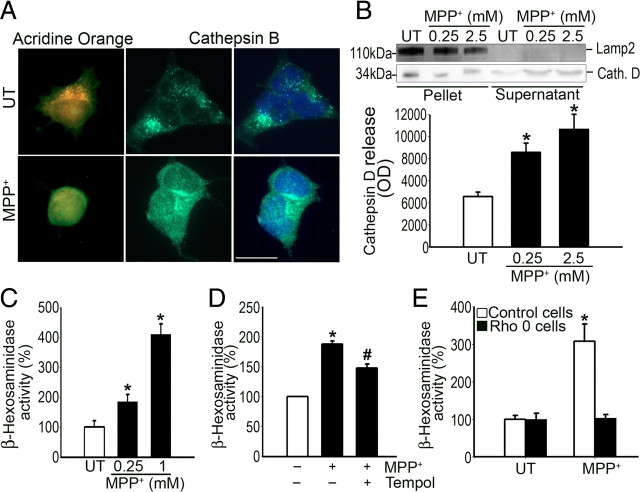
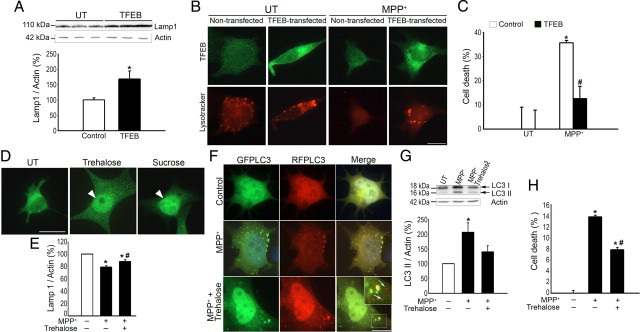
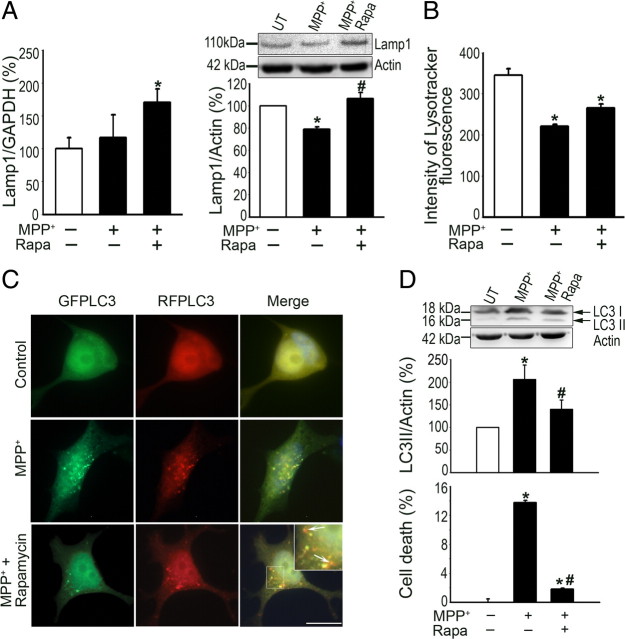
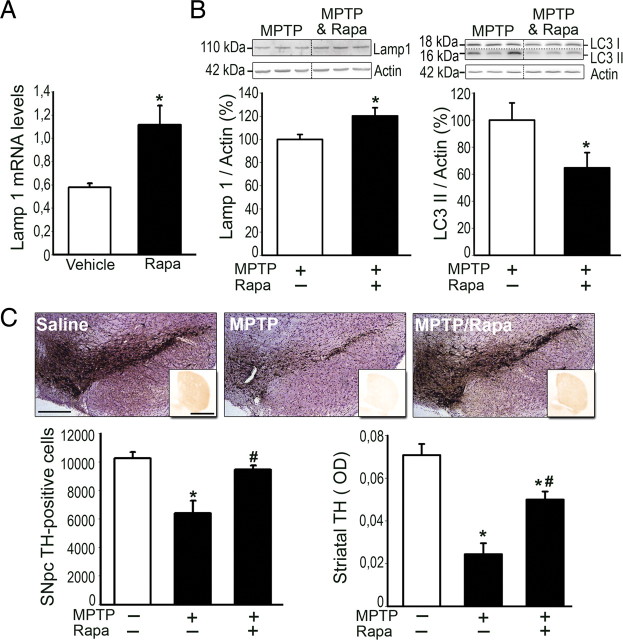
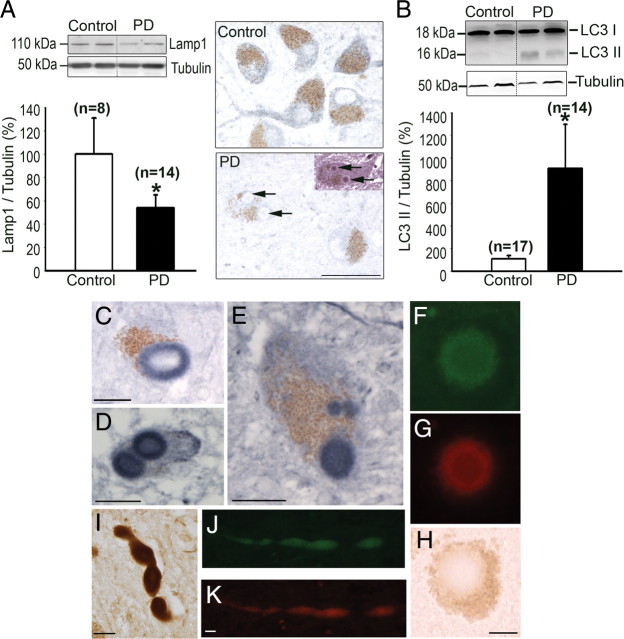
Mounting evidence suggests a role for autophagy dysregulation in Parkinson's disease (PD). The bulk degradation of cytoplasmic proteins (including α-synuclein) and organelles (such as mitochondria) is mediated by macroautophagy, which involves the sequestration of cytosolic components into autophagosomes (AP) and its delivery to lysosomes. Accumulation of AP occurs in postmortem brain samples from PD patients, which has been widely attributed to an induction of autophagy. However, the cause and pathogenic significance of these changes remain unknown. Here we found in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine mouse model of PD that AP accumulation and dopaminergic cell death are preceded by a marked decrease in the amount of lysosomes within dopaminergic neurons. Lysosomal depletion was secondary to the abnormal permeabilization of lysosomal membranes induced by increased mitochondrial-derived reactive oxygen species. Lysosomal permeabilization resulted in a defective clearance and subsequent accumulation of undegraded AP and contributed directly to neurodegeneration by the ectopic release of lysosomal proteases into the cytosol. Lysosomal breakdown and AP accumulation also occurred in PD brain samples, where Lewy bodies were strongly immunoreactive for AP markers. Induction of lysosomal biogenesis by genetic or pharmacological activation of lysosomal transcription factor EB restored lysosomal levels, increased AP clearance and attenuated 1-methyl-4-phenylpyridinium-induced cell death. Similarly, the autophagy-enhancer compound rapamycin attenuated PD-related dopaminergic neurodegeneration, both in vitro and in vivo, by restoring lysosomal levels. Our results indicate that AP accumulation in PD results from defective lysosomal-mediated AP clearance secondary to lysosomal depletion. Restoration of lysosomal levels and function may thus represent a novel neuroprotective strategy in PD.
Figures
References
-
- Aharon-Peretz J, Rosenbaum H, Gershoni-Baruch R. Mutations in the glucocerebrosidase gene and Parkinson's disease in Ashkenazi Jews. N Engl J Med. 2004;351:1972–1977. - PubMed
-
- Anglade P, Vyas S, Javoy-Agid F, Herrero MT, Michel PP, Marquez J, Mouatt-Prigent A, Ruberg M, Hirsch EC, Agid Y. Apoptosis and autophagy in nigral neurons of patients with Parkinson's disease. Histol Histopathol. 1997;12:25–31. - PubMed
-
- Barrachina M, Maes T, Buesa C, Ferrer I. Lysosome-associated membrane protein 1 (LAMP-1) in Alzheimer's disease. Neuropathol Appl Neurobiol. 2006;32:505–516. - PubMed
-
- Boya P, Kroemer G. Lysosomal membrane permeabilization in cell death. Oncogene. 2008;27:6434–6451. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials