

Development of a nascent galectin-1 chimeric molecule for studying the role of leukocyte galectin-1 ligands and immune disease modulation
- PMID: 20844192
- PMCID: PMC2953643
- DOI: 10.4049/jimmunol.1000715
Development of a nascent galectin-1 chimeric molecule for studying the role of leukocyte galectin-1 ligands and immune disease modulation
Abstract
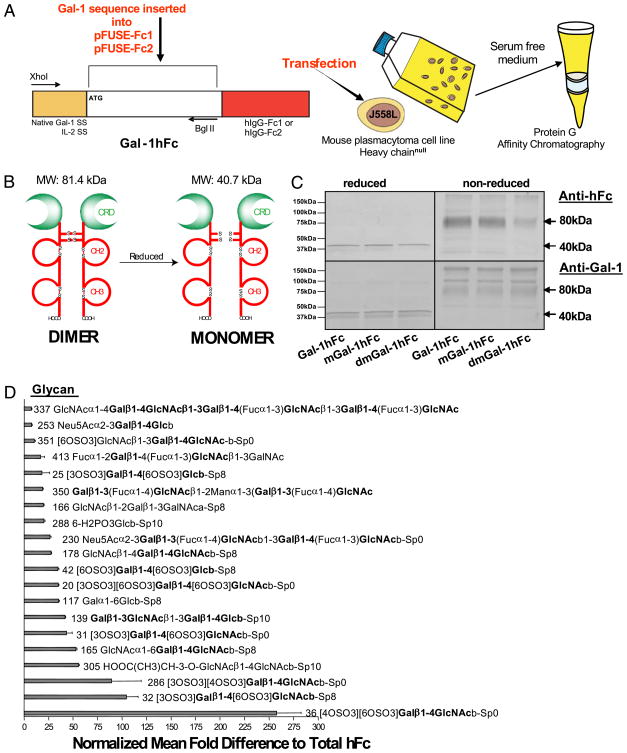
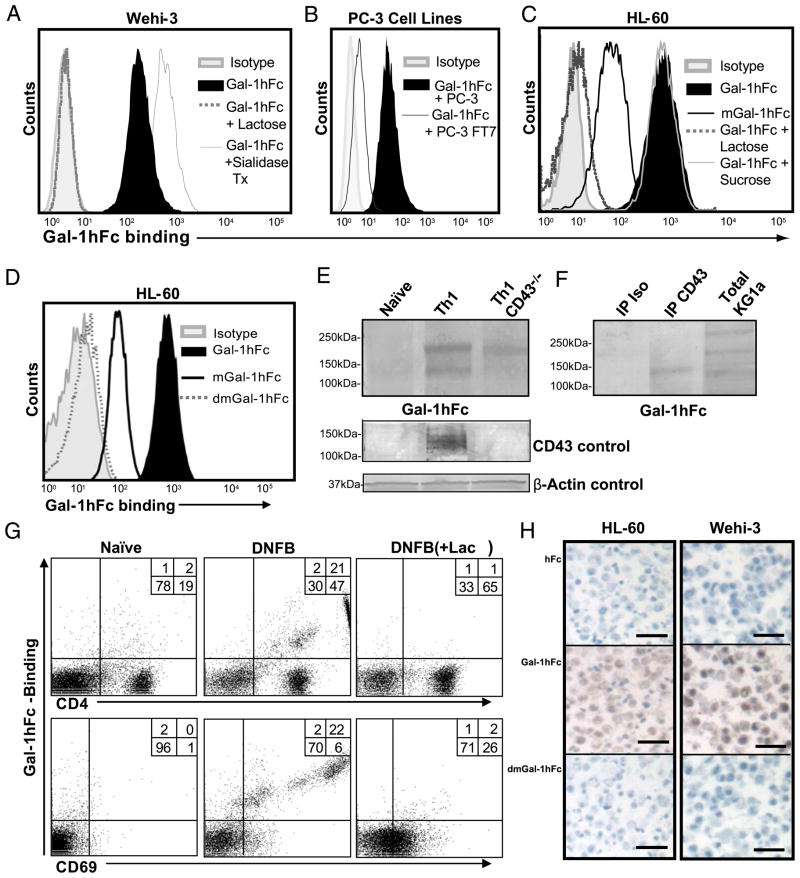
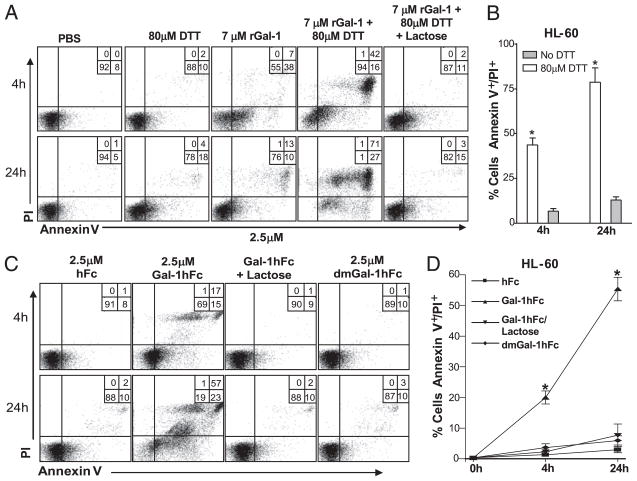
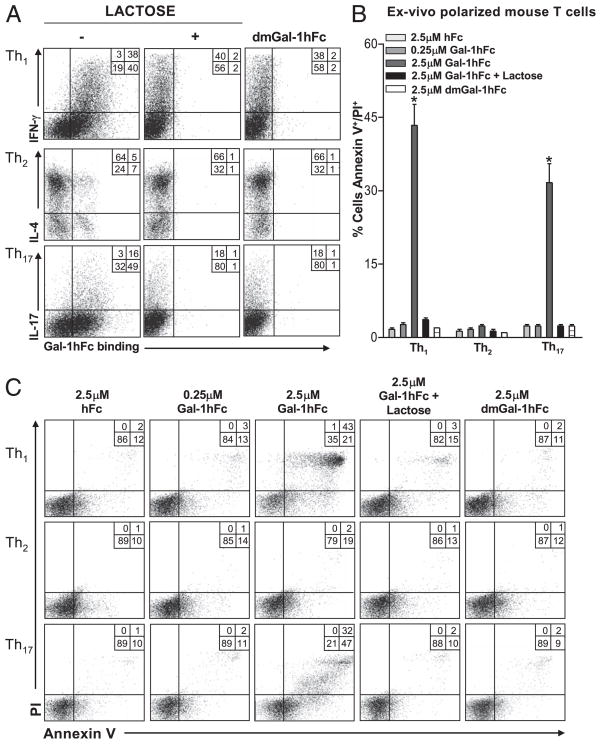
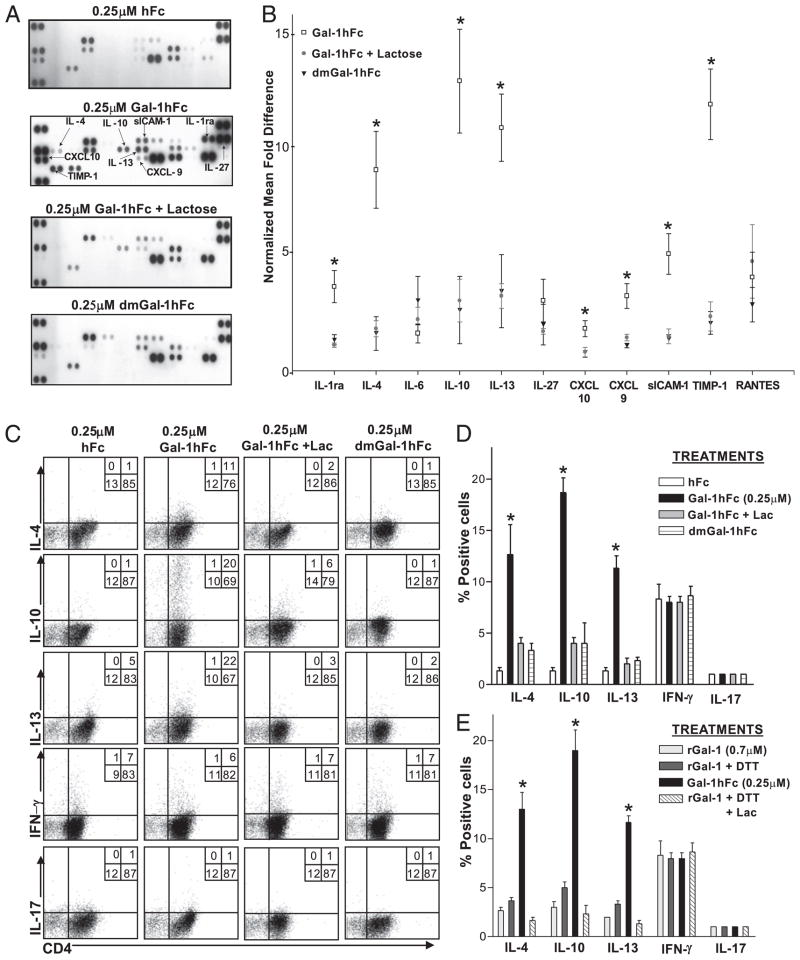
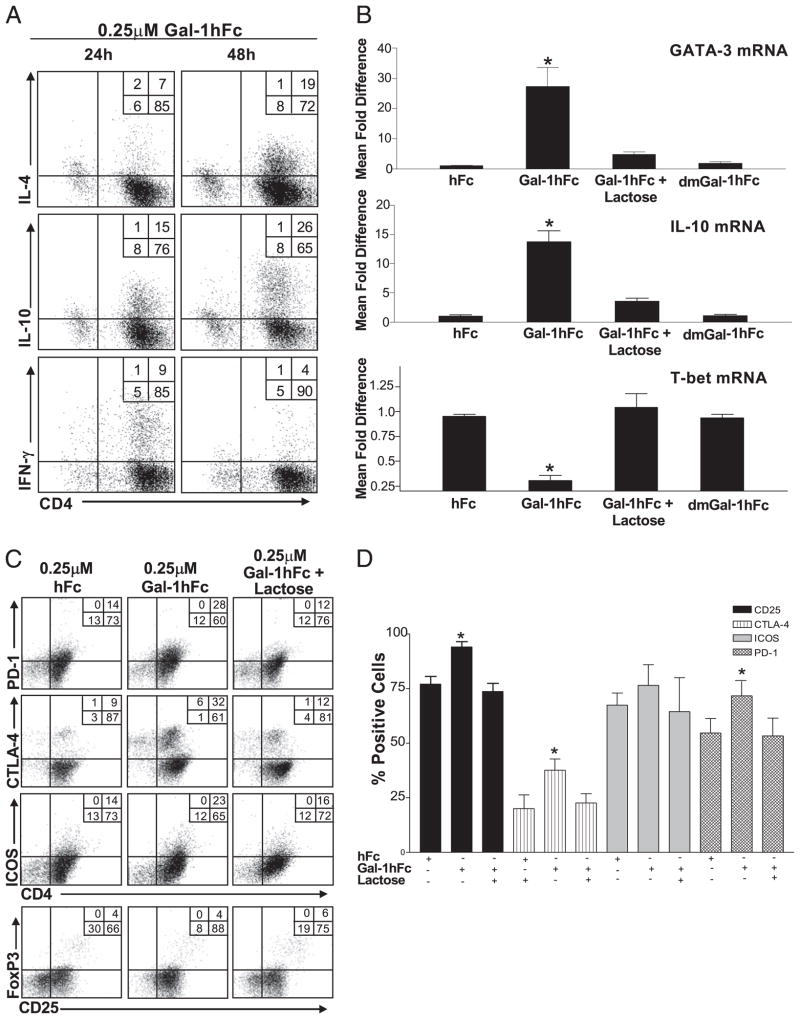
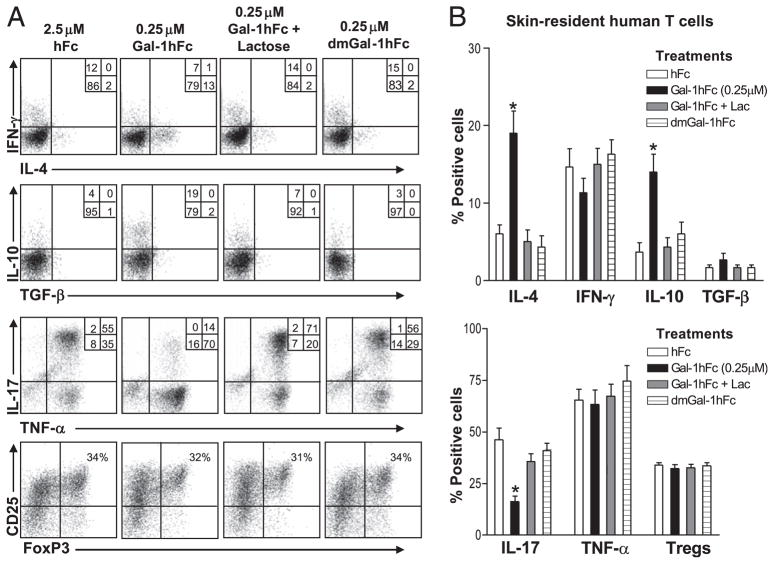
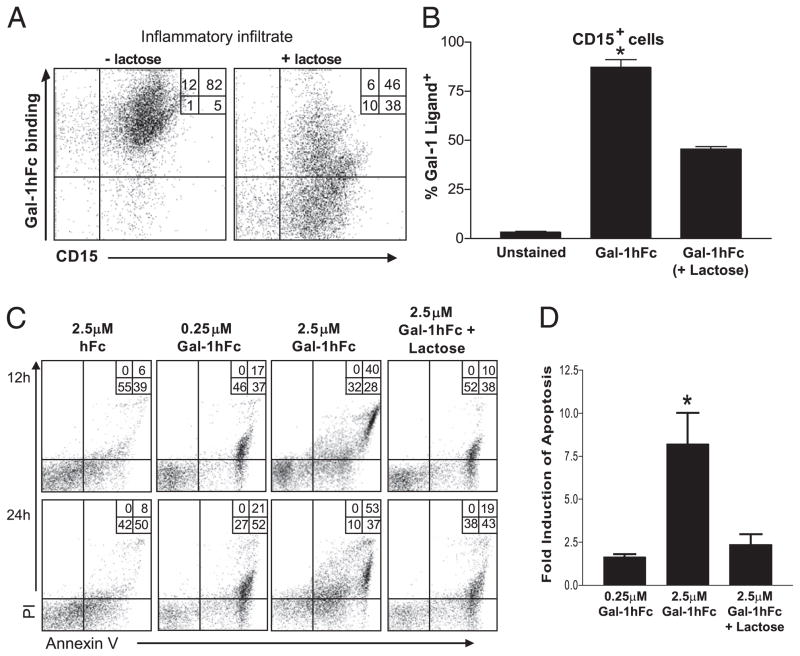
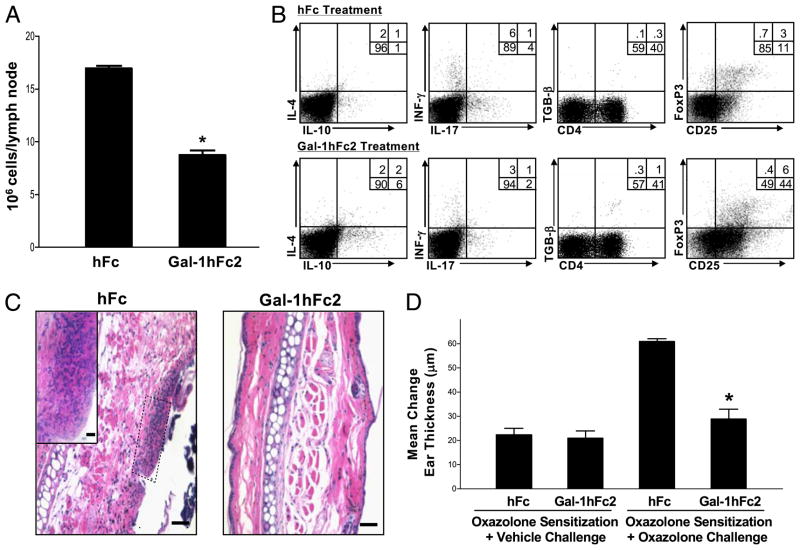
Galectin-1 (Gal-1), a β-galactoside-binding lectin, plays a profound role in modulating adaptive immune responses by altering the phenotype and fate of T cells. Experimental data showing recombinant Gal-1 (rGal-1) efficacy on T cell viability and cytokine production, nevertheless, is controversial due to the necessity of using stabilizing chemicals to help retain Gal-1 structure and function. To address this drawback, we developed a mouse Gal-1 human Ig chimera (Gal-1hFc) that did not need chemical stabilization for Gal-1 ligand recognition, apoptosis induction, and cytokine modulation in a variety of leukocyte models. At high concentrations, Gal-1hFc induced apoptosis in Gal-1 ligand(+) Th1 and Th17 cells, leukemic cells, and granulocytes from synovial fluids of patients with rheumatoid arthritis. Importantly, at low, more physiologic concentrations, Gal-1hFc retained its homodimeric form without losing functionality. Not only did Gal-1hFc-binding trigger IL-10 and Th2 cytokine expression in activated T cells, but members of the CD28 family and several other immunomodulatory molecules were upregulated. In a mouse model of contact hypersensitivity, we found that a non-Fc receptor-binding isoform of Gal-1hFc, Gal-1hFc2, alleviated T cell-dependent inflammation by increasing IL-4(+), IL-10(+), TGF-β(+), and CD25(high)/FoxP3(+) T cells, and by decreasing IFN-γ(+) and IL-17(+) T cells. Moreover, in human skin-resident T cell cultures, Gal-1hFc diminished IL-17(+) T cells and increased IL-4(+) and IL-10(+) T cells. Gal-1hFc will not only be a useful new tool for investigating the role of Gal-1 ligands in leukocyte death and cytokine stimulation, but for studying how Gal-1-Gal-1 ligand binding shapes the intensity of immune responses.
Figures
References
-
- Ilarregui JM, Croci DO, Bianco GA, Toscano MA, Salatino M, Vermeulen ME, Geffner JR, Rabinovich GA. Tolerogenic signals delivered by dendritic cells to T cells through a galectin-1-driven immunoregulatory circuit involving interleukin 27 and interleukin 10. Nat Immunol. 2009;10:981–991. - PubMed
-
- Garín MI, Chu CC, Golshayan D, Cernuda-Morollón E, Wait R, Lechler RI. Galectin-1: a key effector of regulation mediated by CD4+ CD25+ T cells. Blood. 2007;109:2058–2065. - PubMed
-
- Toscano MA, Bianco GA, Ilarregui JM, Croci DO, Correale J, Hernandez JD, Zwirner NW, Poirier F, Riley EM, Baum LG, Rabinovich GA. Differential glycosylation of TH1, TH2 and TH-17 effector cells selectively regulates susceptibility to cell death. Nat Immunol. 2007;8:825–834. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
