

Role of epoxyeicosatrienoic acids as autocrine metabolites in glutamate-mediated K+ signaling in perivascular astrocytes
- PMID: 20844244
- PMCID: PMC2980321
- DOI: 10.1152/ajpcell.00225.2010
Role of epoxyeicosatrienoic acids as autocrine metabolites in glutamate-mediated K+ signaling in perivascular astrocytes
Abstract
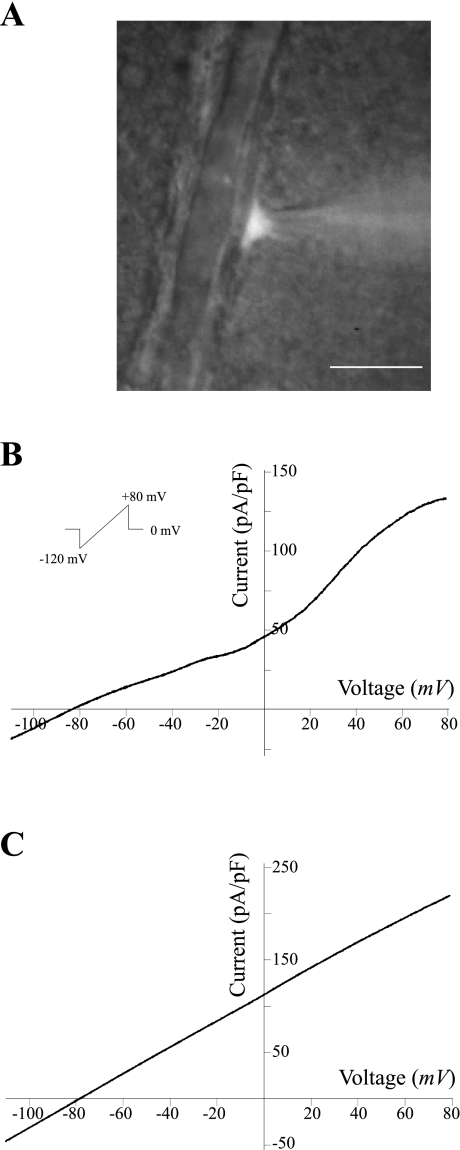
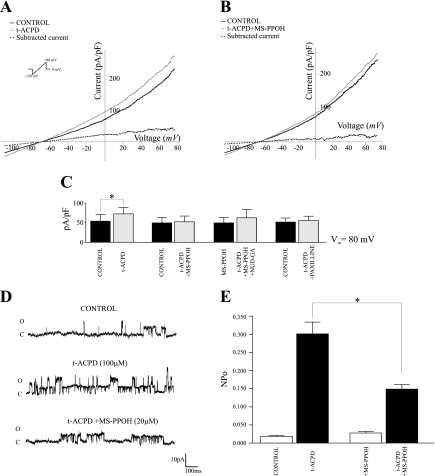
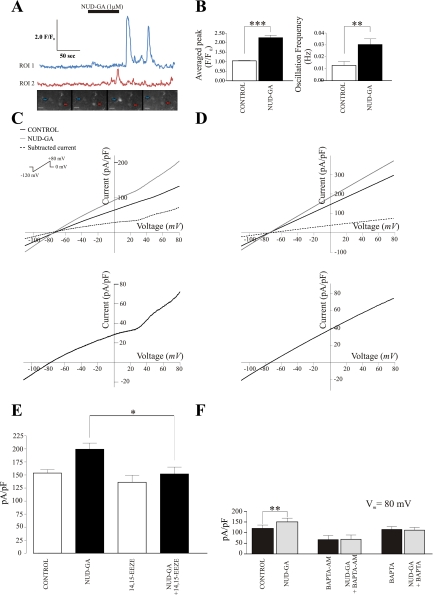
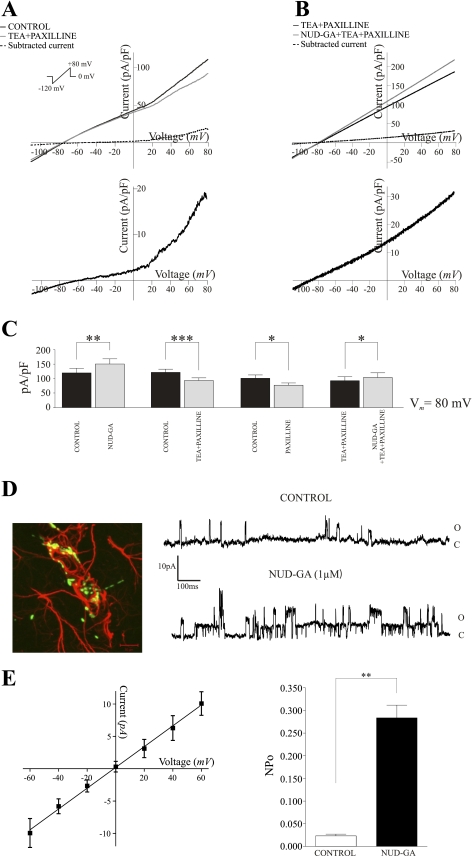
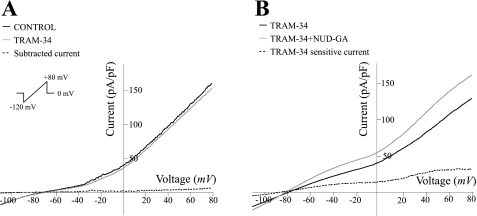
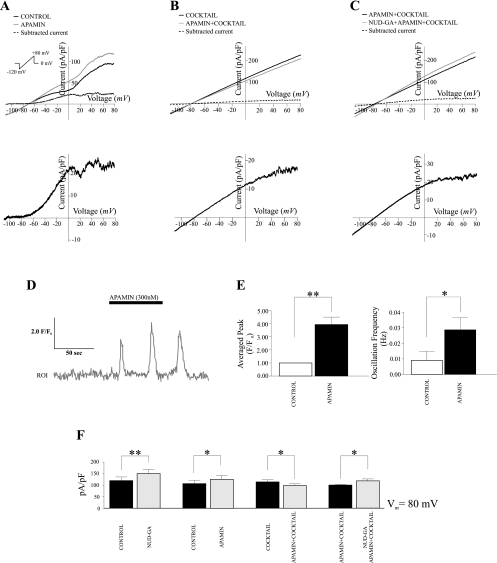
Epoxyeicosatrienoic acids (EETs), synthesized and released by astrocytes in response to glutamate, are known to play a pivotal role in neurovascular coupling. In vascular smooth muscle cells (VSMC), EETs activate large-conductance, Ca(2+)-activated K(+) (BK) channels resulting in hyperpolarization and vasodilation. However, the functional role and mechanism of action for glial-derived EETs are still to be determined. In this study, we evaluated the effect of the synthetic EET analog 11-nonyloxy-undec-8(Z)-enoic acid (NUD-GA) on outward K(+) currents mediated by calcium-activated K(+) channels. Addition of NUD-GA significantly increased intracellular Ca(2+) and outward K(+) currents in perivascular astrocytes. NUD-GA-induced currents were significantly inhibited by BK channel blockers paxilline and tetraethylammonium (TEA) (23.4 ± 2.4%; P < 0.0005). Similarly, NUD-GA-induced currents were also significantly inhibited in the presence of the small-conductance Ca(2+)-activated K(+) channel inhibitor apamin along with a combination of blockers against glutamate receptors (12.8 ± 2.70%; P < 0.05). No changes in outward currents were observed in the presence of the channel blocker for intermediate-conductance K(+) channels TRAM-34. Blockade of the endogenous production of EETs with N-methylsulfonyl-6-(2-propargyloxyphenyl)hexanamide (MS-PPOH) significantly blunted (dl)-1-aminocyclopentane-trans-1,3-dicarboxylic acid (t-ACPD)-induced outward K(+) currents (P < 0.05; n = 6). Both NUD-GA and t-ACPD significantly increased BK channel single open probability; the later was blocked following MS-PPOH incubation. Our data supports the idea that EETs are potent K(+) channel modulators in cortical perivascular astrocytes and further suggest that these metabolites may participate in NVC by modulating the levels of K(+) released at the gliovascular space.
Figures
References
-
- Alkayed NJ, Birks EK, Narayanan J, Petrie KA, Kohler-Cabot AE, Harder DR. Role of P-450 arachidonic acid epoxygenase in the response of cerebral blood flow to glutamate in rats. Stroke 28: 1066–1072, 1997 - PubMed
-
- Alkayed NJ, Narayanan J, Gebremedhin D, Medhora M, Roman RJ, Harder DR. Molecular characterization of an arachidonic acid epoxygenase in rat brain astrocytes. Stroke 27: 971–979, 1996 - PubMed
-
- Amruthesh SC, Falck JR, Ellis EF. Brain synthesis and cerebrovascular action of epoxygenase metabolites of arachidonic acid. J Neurochem 58: 503–510, 1992 - PubMed
-
- Armstrong WE, Rubrum A, Teruyama R, Bond CT, Adelman JP. Immunocytochemical localization of small-conductance, calcium-dependent potassium channels in astrocytes of the rat supraoptic nucleus. J Comp Neurol 491: 175–185, 2005 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous