

Syntaxin 3 is necessary for cAMP- and cGMP-regulated exocytosis of CFTR: implications for enterotoxigenic diarrhea
- PMID: 20844248
- PMCID: PMC3006332
- DOI: 10.1152/ajpcell.00029.2010
Syntaxin 3 is necessary for cAMP- and cGMP-regulated exocytosis of CFTR: implications for enterotoxigenic diarrhea
Abstract
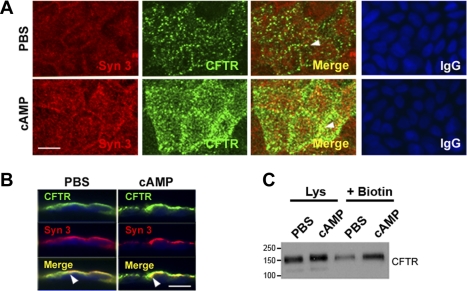
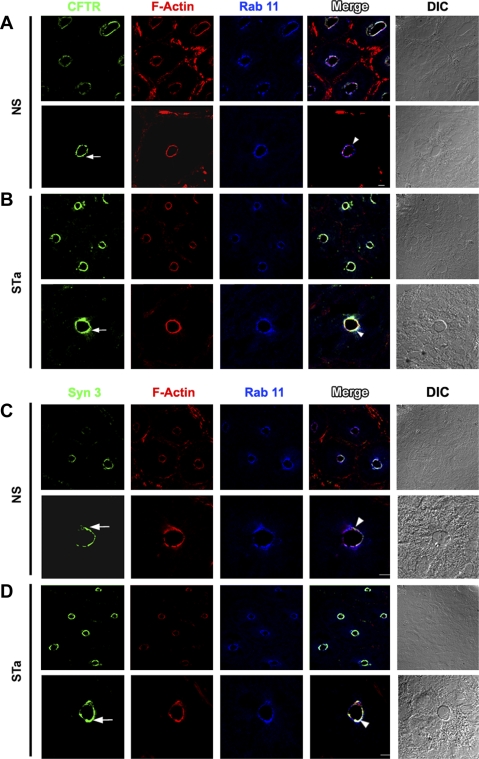
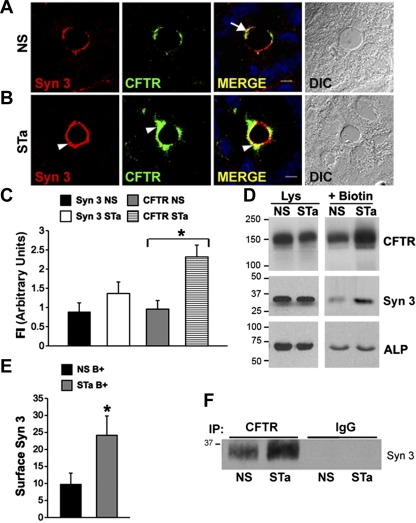
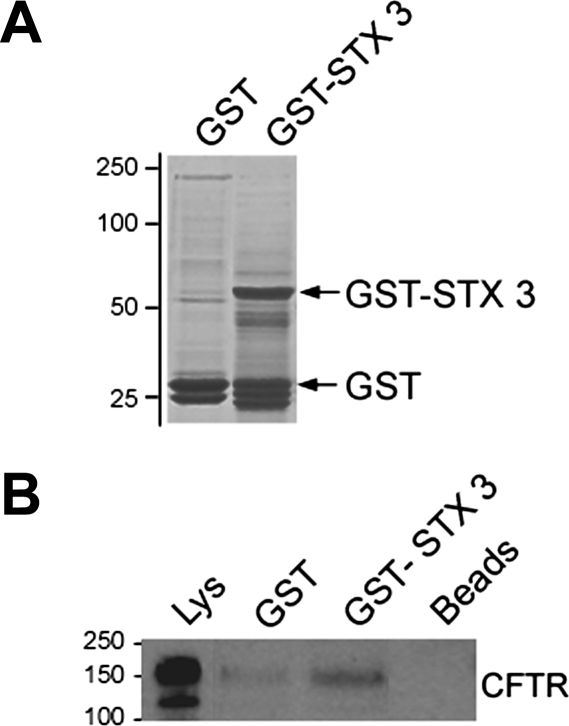
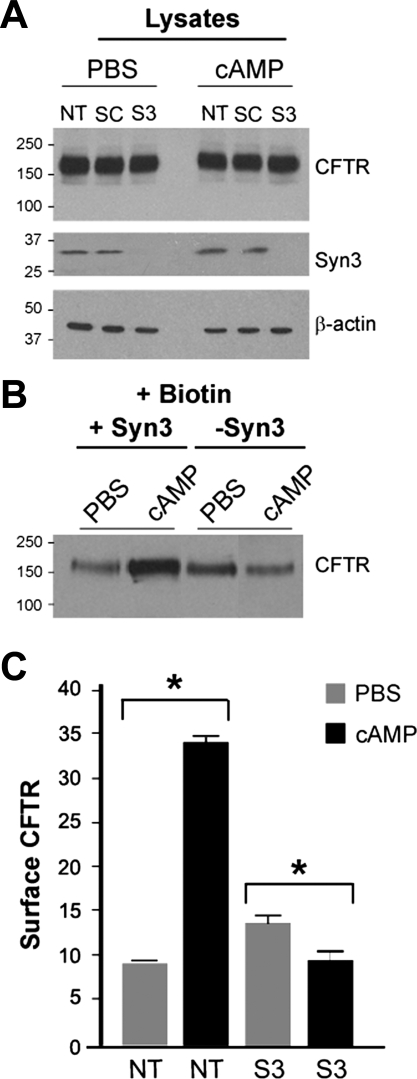
Enterotoxins elaborated by Vibrio cholerae and Escherichia coli cannot elicit fluid secretion in the absence of functional cystic fibrosis transmembrane conductance regulator (CFTR) chloride channels. After enterotoxin exposure, CFTR channels are rapidly recruited from endosomes and undergo exocytic insertion into the apical plasma membrane of enterocytes to increase the number of channels on the cell surface by at least fourfold. However, the molecular machinery that orchestrates exocytic insertion of CFTR into the plasma membrane is largely unknown. The present study used immunofluorescence, immunoblotting, surface biotinylation, glutathione S-transferase (GST) pulldown assays, and immunoprecipitation to identify components of the exocytic soluble N-ethylmaleimide (NEM)-sensitive factor attachment receptor (SNARE) vesicle fusion machinery in cyclic nucleotide-activated exocytosis of CFTR in rat jejunum and polarized intestinal Caco-2(BB)e cells. Syntaxin 3, an intestine-specific SNARE, colocalized with CFTR on the apical domain of enterocytes in rat jejunum and polarized Caco-2(BB)e cells. Coimmunoprecipitation and GST binding studies confirmed that syntaxin 3 interacts with CFTR in vivo. Moreover, heat-stable enterotoxin (STa) activated exocytosis of both CFTR and syntaxin 3 to the surface of rat jejunum. Silencing of syntaxin 3 by short hairpin RNA (shRNA) interference abrogated cyclic nucleotide-stimulated exocytosis of CFTR in cells. These observations reveal a new and important role for syntaxin 3 in the pathophysiology of enterotoxin-elicited diarrhea.
Figures
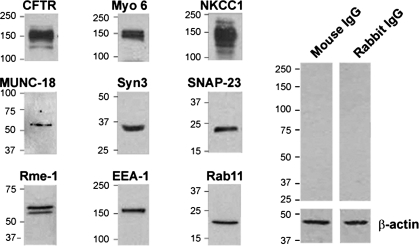
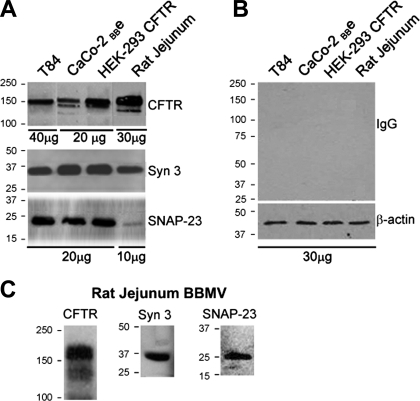
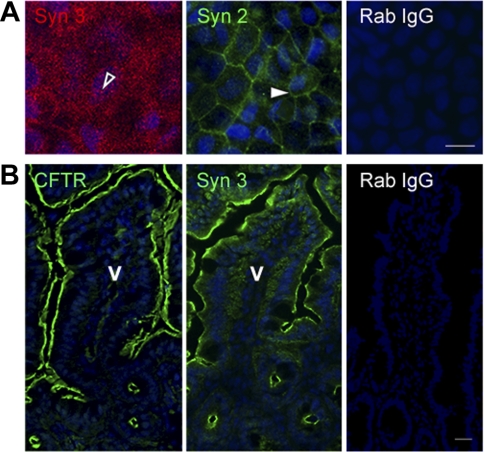
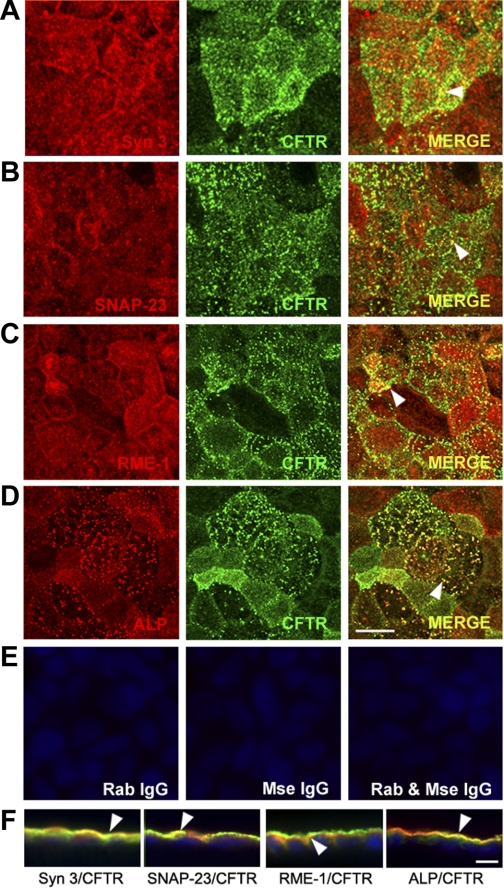
References
-
- Ameen N, Alexis J, Salas P. Cellular localization of the cystic fibrosis transmembrane conductance regulator in mouse intestinal tract. Histochem Cell Biol 114: 69– 75, 2000 - PubMed
-
- Ameen N, Apodaca G. Defective CFTR apical endocytosis and enterocyte brush border in myosin VI-deficient mice. Traffic 8: 998– 1006, 2007 - PubMed
-
- Ameen NA, Marino C, Salas PJ. cAMP-dependent exocytosis and vesicle traffic regulate CFTR and fluid transport in rat jejunum in vivo. Am J Physiol Cell Physiol 284: C429– C438, 2003 - PubMed
-
- Ameen NA, Martensson B, Bourguinon L, Marino C, Isenberg J. CFTR channel insertion to the apical surface in rat duodenal villus epithelial cells is upregulated by VIP in vivo. J Cell Sci 112: 887– 894, 1999 - PubMed
-
- Ameen NA, van Donselaar E, Posthuma G, de Jonge H, McLaughlin G, Geuze HJ, Marino C, Peters PJ. Subcellular distribution of CFTR in rat intestine supports a physiologic role for CFTR regulation by vesicle traffic. Histochem Cell Biol 114: 219– 228, 2000 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials