

Nucleo-cytoplasmic localization domains regulate Krüppel-like factor 6 (KLF6) protein stability and tumor suppressor function
- PMID: 20844588
- PMCID: PMC2936564
- DOI: 10.1371/journal.pone.0012639
Nucleo-cytoplasmic localization domains regulate Krüppel-like factor 6 (KLF6) protein stability and tumor suppressor function
Abstract
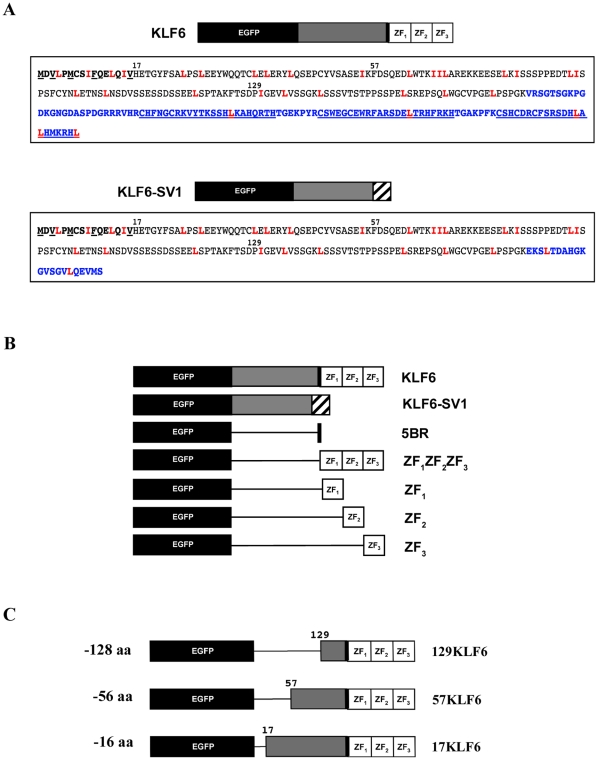
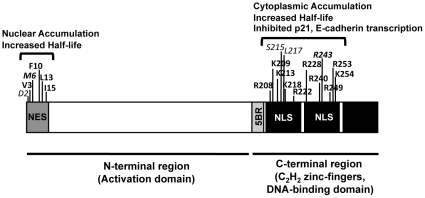
Background: The tumor suppressor KLF6 and its oncogenic cytoplasmic splice variant KLF6-SV1 represent a paradigm in cancer biology in that their antagonistic cancer functions are encoded within the same gene. As a consequence of splicing, KLF6-SV1 loses both the C-terminus C2H2 three zinc finger (ZF) domain, which characterizes all KLF proteins, as well as the adjacent 5' basic region (5BR), a putative nuclear localization signal (NLS). It has been hypothesized that this NLS is a functional domain critical to direct the distinct subcellular localization of the tumor suppressor and its splice variant.
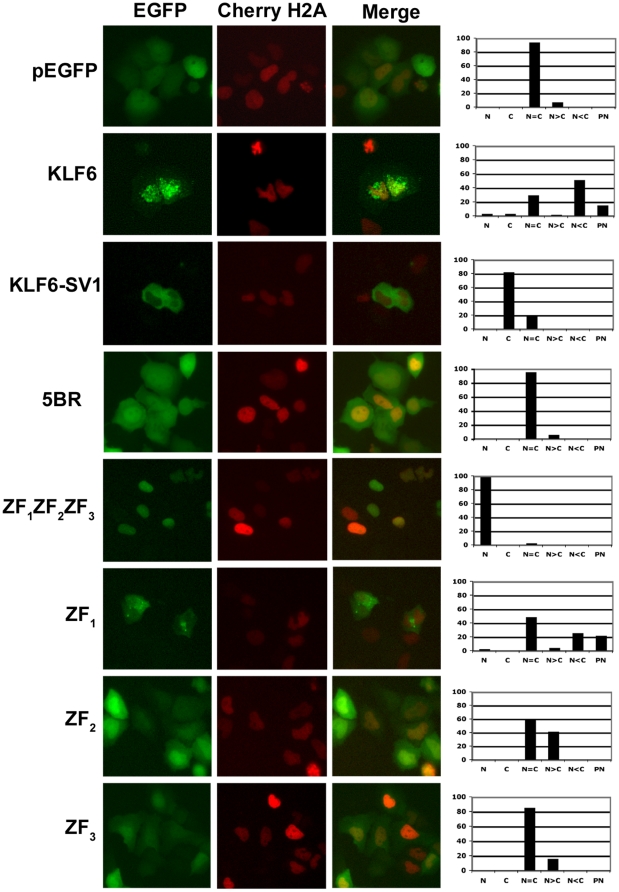
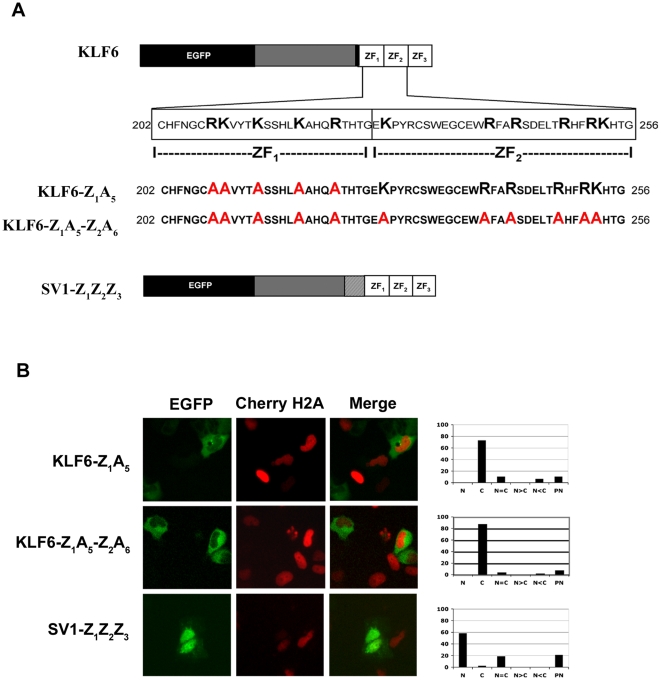
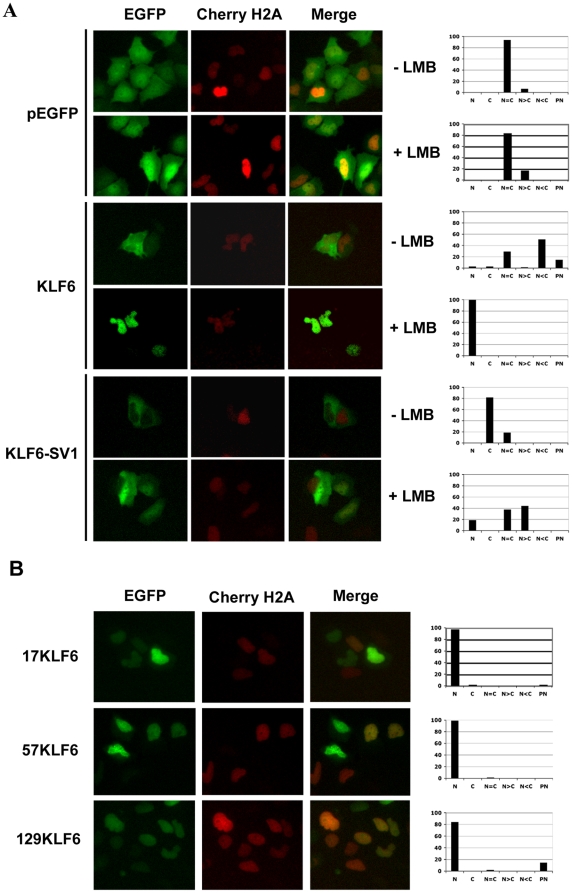
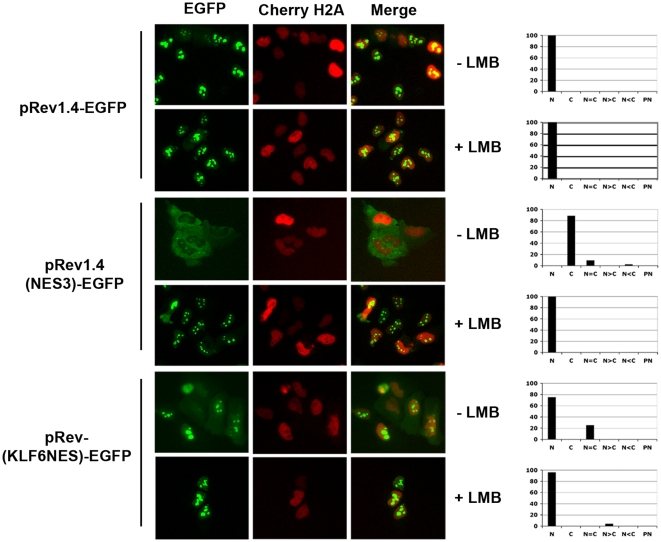
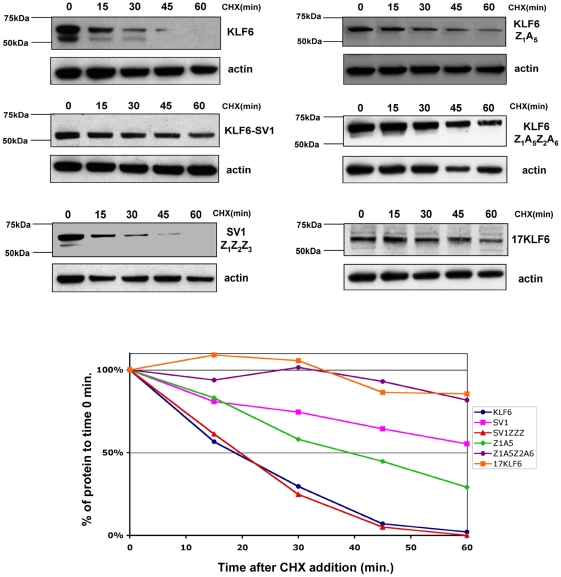
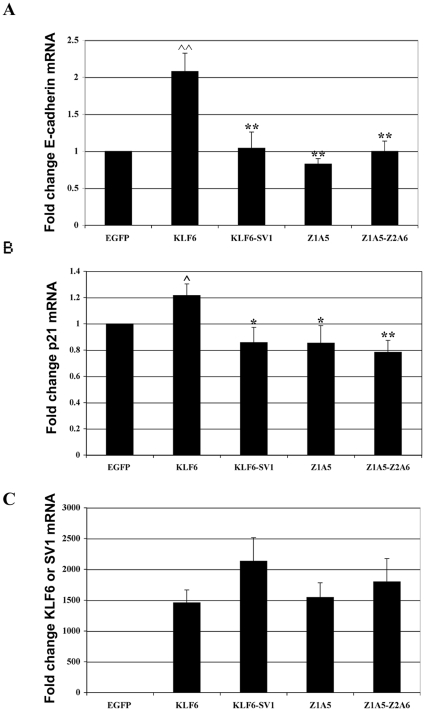
Methodology/principal findings: In this study, we demonstrate using EGFP fusion constructs that KLF6/KLF6-SV1 nucleo-cytoplasmic transport is not regulated by the 5' basic region but activated by a novel NLS encoded within the ZF domain, and a nuclear export signal (NES) located in the first 16 amino acids of the shared N-terminus sequence. We demonstrate KLF6 nuclear export to be Crm1-dependent. The dysregulation of nucleo-cytoplasmic transport when disrupting the KLF6 NLS using site-directed mutagenesis showed that its integrity is necessary for appropriate protein stability. Moreover, these mutations impaired transcriptional induction of two KLF6 well-characterized target genes, E-cadherin and p21, as shown by RT-PCR and luciferase promoter assays. The addition of the ZF domain to KLF6-SV1 results in its nuclear localization and a markedly decreased half-life similar to wild type KLF6.
Conclusions/significance: We describe the domains that control KLF6 nucleo-cytoplasmic shuttling and how these domains play a role in KLF6 protein half-life and tumor suppressor function. The results begin to mechanistically explain, at least in part, the opposing functions of KLF6 and KLF6-SV1 in cancer.
Conflict of interest statement
Figures
References
-
- Narla G, Heath KE, Reeves HL, Li D, Giono LE, et al. KLF6, a candidate tumor suppressor gene mutated in prostate cancer. Science. 2001;294:2563–2566. - PubMed
-
- Black AR, Black JD, Azizkhan-Clifford J. Sp1 and krüppel-like factor family of transcription factors in cell growth regulation and cancer. J Cell Physiol. 2001;188:143–160. - PubMed
-
- DiFeo A, Narla G, Hirshfeld J, Camacho-Vanegas O, Narla J, et al. Roles of KLF6 and KLF6-SV1 in ovarian cancer progression and intraperitoneal dissemination. Clin Cancer Res. 2006;12:3730–3739. - PubMed
-
- Camacho-Vanegas O, Narla G, Teixeira MS, DiFeo A, Misra A, et al. Functional inactivation of the KLF6 tumor suppressor gene by loss of heterozygosity and increased alternative splicing in glioblastoma. Int J Cancer. 2007;121:1390–1395. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
