

Mutational and biochemical analysis of the DNA-entry nuclease EndA from Streptococcus pneumoniae
- PMID: 20846957
- PMCID: PMC3025545
- DOI: 10.1093/nar/gkq802
Mutational and biochemical analysis of the DNA-entry nuclease EndA from Streptococcus pneumoniae
Abstract
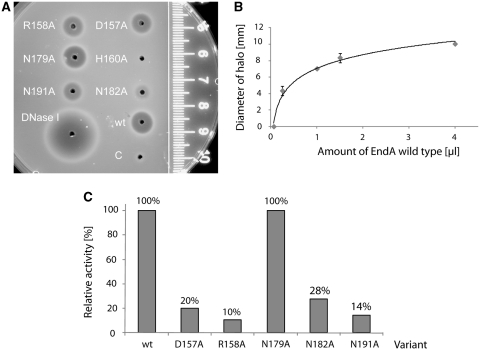
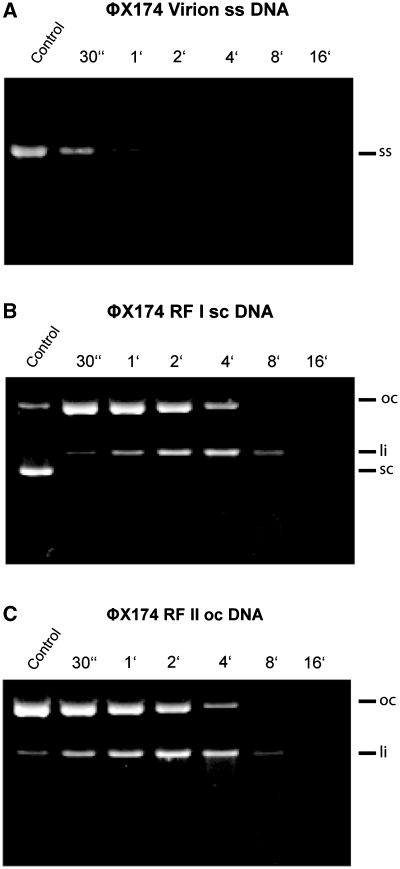
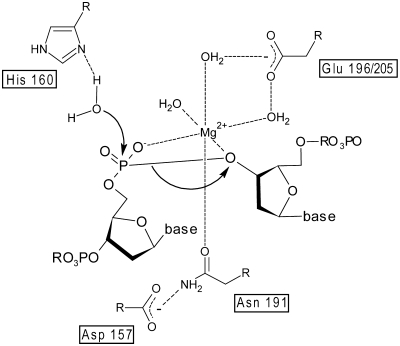
EndA is a membrane-attached surface-exposed DNA-entry nuclease previously known to be required for genetic transformation of Streptococcus pneumoniae. More recent studies have shown that the enzyme also plays an important role during the establishment of invasive infections by degrading extracellular chromatin in the form of neutrophil extracellular traps (NETs), enabling streptococci to overcome the innate immune system in mammals. As a virulence factor, EndA has become an interesting target for future drug design. Here we present the first mutational and biochemical analysis of recombinant forms of EndA produced either in a cell-free expression system or in Escherichia coli. We identify His160 and Asn191 to be essential for catalysis and Asn182 to be required for stability of EndA. The role of His160 as the putative general base in the catalytic mechanism is supported by chemical rescue of the H160A variant of EndA with imidazole added in excess. Our study paves the way for the identification and development of protein or low-molecular-weight inhibitors for EndA in future high-throughput screening assays.
Figures
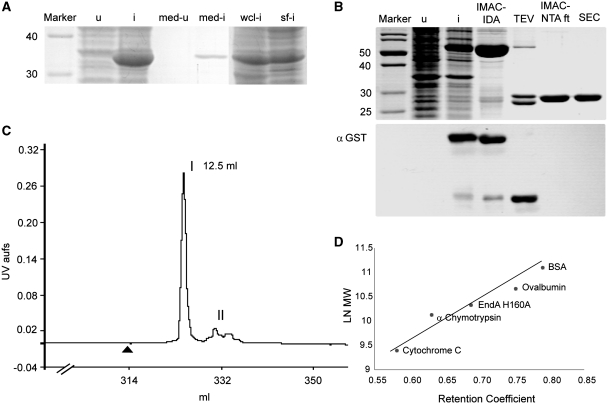
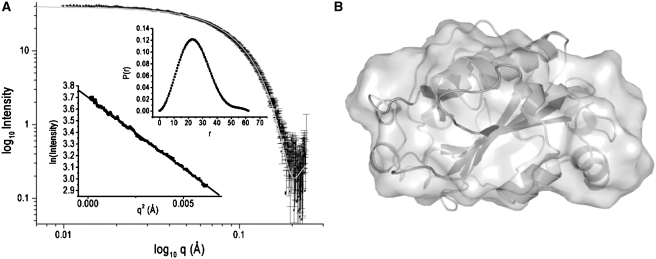
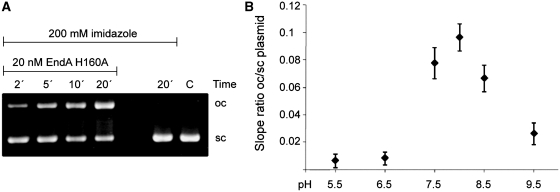
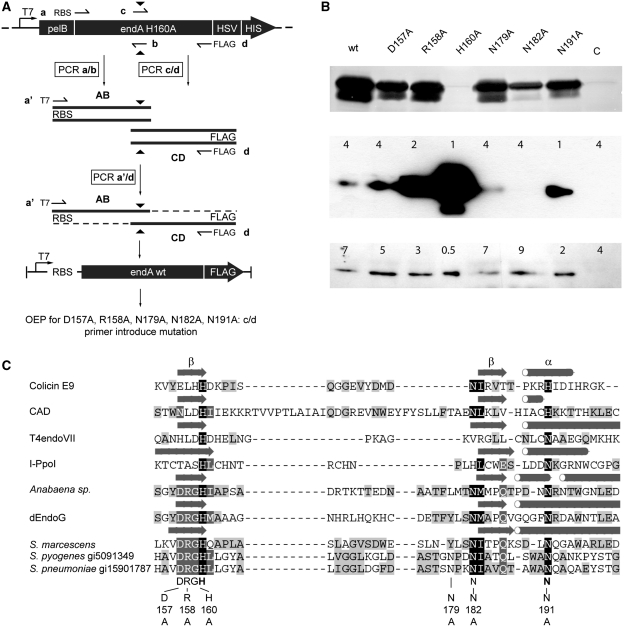
References
-
- Puyet A, Greenberg B, Lacks SA. Genetic and structural characterization of endA. A membrane-bound nuclease required for transformation of Streptococcus pneumoniae. J. Mol. Biol. 1990;213:727–738. - PubMed
-
- Buchanan JT, Simpson AJ, Aziz RK, Liu GY, Kristian SA, Kotb M, Feramisco J, Nizet V. DNase expression allows the pathogen group A Streptococcus to escape killing in neutrophil extracellular traps. Curr. Biol. 2006;16:396–400. - PubMed
-
- Beiter K, Wartha F, Albiger B, Normark S, Zychlinsky A, Henriques-Normark B. An endonuclease allows Streptococcus pneumoniae to escape from neutrophil extracellular traps. Curr. Biol. 2006;16:401–407. - PubMed
-
- Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, Weinrauch Y, Zychlinsky A. Neutrophil extracellular traps kill bacteria. Science. 2004;303:1532–1535. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
