

Regulation of replication termination by Reb1 protein-mediated action at a distance
- PMID: 20850009
- PMCID: PMC2945231
- DOI: 10.1016/j.cell.2010.08.013
Regulation of replication termination by Reb1 protein-mediated action at a distance
Abstract
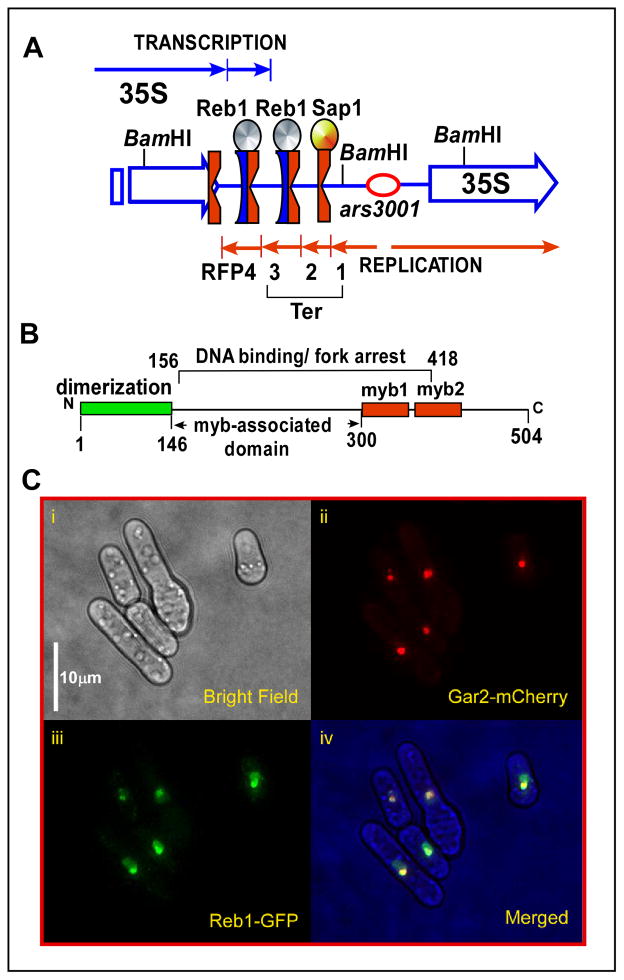
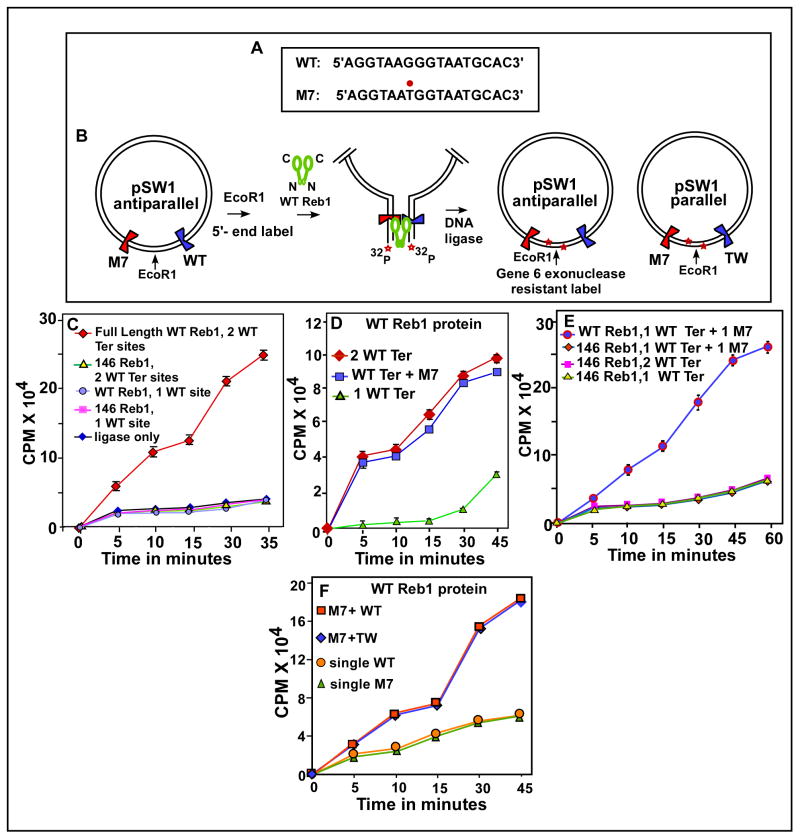
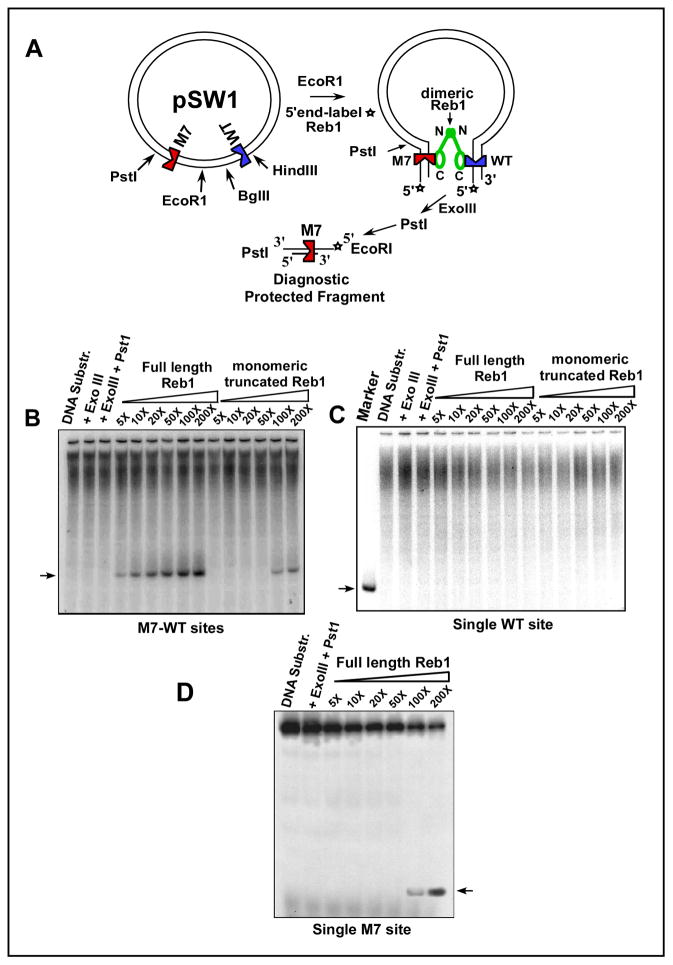
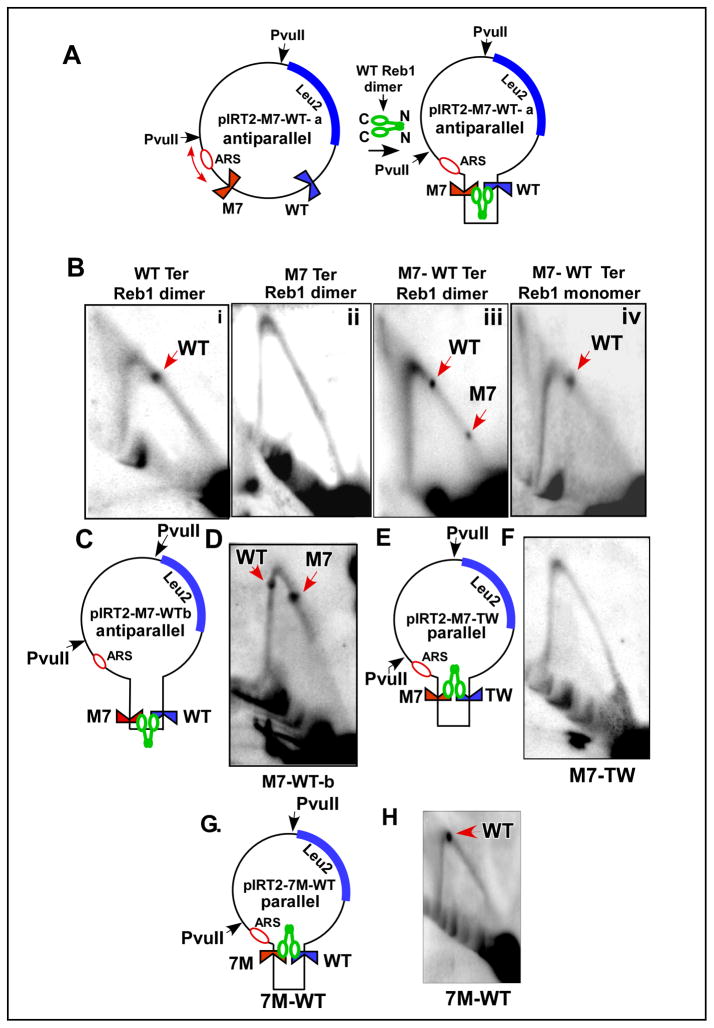
DNA transactions driven by long-range protein-mediated inter- and intrachromosomal interactions have been reported to influence gene expression. Here, we report that site-specific replication termination in Schizosaccharomyces pombe is modulated by protein-mediated interactions between pairs of Ter sites located either on the same or on different chromosomes. The dimeric Reb1 protein catalyzes termination and mediates interaction between Ter sites. The Reb1-dependent interactions between two antiparallel Ter sites in cis caused looping out of the intervening DNA in vitro and enhancement of fork arrest in vivo. A Ter site on chromosome 2 interacted pairwise with two Ter sites located on chromosome 1 by chromosome kissing. Mutational inactivation of the major interacting Ter site on chromosome 1 significantly reduced fork arrest at the Ter site on chromosome 2, thereby revealing a cooperative mechanism of control of replication termination.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
Similar articles
-
Mechanistic insights into replication termination as revealed by investigations of the Reb1-Ter3 complex of Schizosaccharomyces pombe.Mol Cell Biol. 2008 Nov;28(22):6844-57. doi: 10.1128/MCB.01235-08. Epub 2008 Sep 15. Mol Cell Biol. 2008. PMID: 18794373 Free PMC article.
-
Functional architecture of the Reb1-Ter complex of Schizosaccharomyces pombe.Proc Natl Acad Sci U S A. 2016 Apr 19;113(16):E2267-76. doi: 10.1073/pnas.1525465113. Epub 2016 Mar 28. Proc Natl Acad Sci U S A. 2016. PMID: 27035982 Free PMC article.
-
Crystallization and preliminary X-ray characterization of the eukaryotic replication terminator Reb1-Ter DNA complex.Acta Crystallogr F Struct Biol Commun. 2015 Apr;71(Pt 4):414-8. doi: 10.1107/S2053230X15004112. Epub 2015 Mar 20. Acta Crystallogr F Struct Biol Commun. 2015. PMID: 25849502 Free PMC article.
-
Tus-Ter as a tool to study site-specific DNA replication perturbation in eukaryotes.Cell Cycle. 2014;13(19):2994-8. doi: 10.4161/15384101.2014.958912. Cell Cycle. 2014. PMID: 25486560 Free PMC article. Review.
-
Regulation of chromosome dynamics by Hsk1/Cdc7 kinase.Biochem Soc Trans. 2013 Dec;41(6):1712-9. doi: 10.1042/BST20130217. Biochem Soc Trans. 2013. PMID: 24256280 Review.
Cited by
-
Mechanism and physiological significance of programmed replication termination.Semin Cell Dev Biol. 2014 Jun;30:165-73. doi: 10.1016/j.semcdb.2014.04.030. Epub 2014 May 6. Semin Cell Dev Biol. 2014. PMID: 24811316 Free PMC article. Review.
-
Chromosome domain architecture and dynamic organization of the fission yeast genome.FEBS Lett. 2015 Oct 7;589(20 Pt A):2975-86. doi: 10.1016/j.febslet.2015.06.008. Epub 2015 Jun 19. FEBS Lett. 2015. PMID: 26096785 Free PMC article. Review.
-
Chromosome I controls chromosome II replication in Vibrio cholerae.PLoS Genet. 2014 Feb 27;10(2):e1004184. doi: 10.1371/journal.pgen.1004184. eCollection 2014 Feb. PLoS Genet. 2014. PMID: 24586205 Free PMC article.
-
Regulation of DNA Replication through Natural Impediments in the Eukaryotic Genome.Genes (Basel). 2017 Mar 7;8(3):98. doi: 10.3390/genes8030098. Genes (Basel). 2017. PMID: 28272375 Free PMC article. Review.
-
Impediments to replication fork movement: stabilisation, reactivation and genome instability.Chromosoma. 2013 Mar;122(1-2):33-45. doi: 10.1007/s00412-013-0398-9. Epub 2013 Feb 28. Chromosoma. 2013. PMID: 23446515 Review.
References
-
- Bastia D, Mohanty BK. Mechanisms for completing DNA replication. In: DePamphilis M, editor. DNA Replication in Eukaryotic Cells. Plainview, NY: Cold Spring harbor Laboratory Press; 1996. pp. 177–215.
-
- Bastia D, Mohanty BK. Termination of DNA Replication. In: DePamphilis M, editor. DNA replication and human disease. Cold Spring Harbor, New York: Cold Spring Harbor Laboratory Press; 2006. pp. 155–174.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
