

Structure-based analysis of Toxoplasma gondii profilin: a parasite-specific motif is required for recognition by Toll-like receptor 11
- PMID: 20851125
- PMCID: PMC2957522
- DOI: 10.1016/j.jmb.2010.09.022
Structure-based analysis of Toxoplasma gondii profilin: a parasite-specific motif is required for recognition by Toll-like receptor 11
Abstract
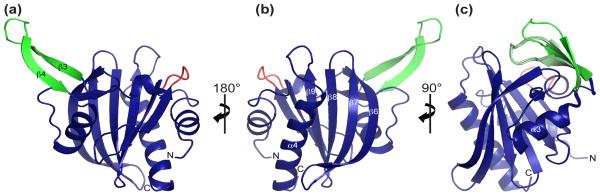
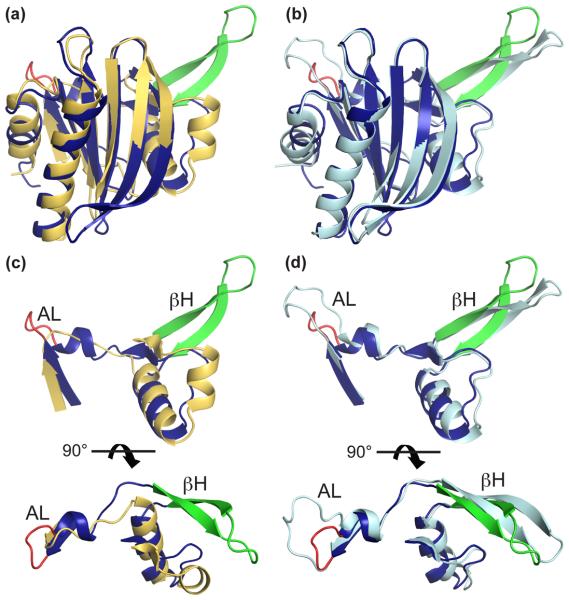
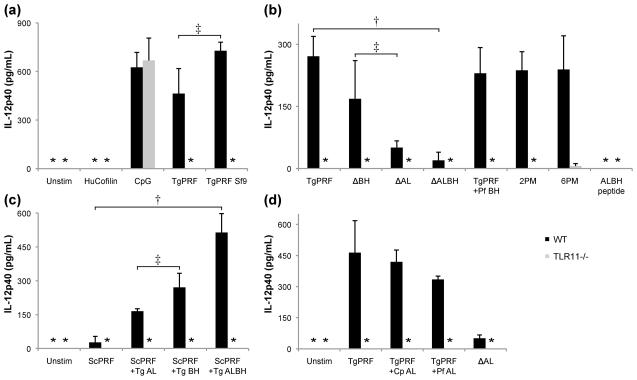
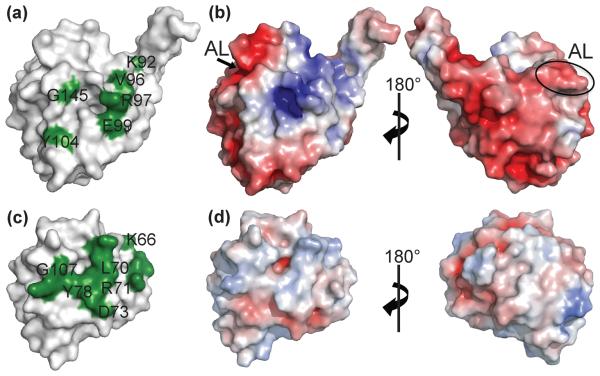
Profilins promote actin polymerization by exchanging ADP for ATP on monomeric actin and delivering ATP-actin to growing filament barbed ends. Apicomplexan protozoa such as Toxoplasma gondii invade host cells using an actin-dependent gliding motility. Toll-like receptor (TLR) 11 generates an innate immune response upon sensing T. gondii profilin (TgPRF). The crystal structure of TgPRF reveals a parasite-specific surface motif consisting of an acidic loop, followed by a long β-hairpin. A series of structure-based profilin mutants show that TLR11 recognition of the acidic loop is responsible for most of the interleukin (IL)-12 secretion response to TgPRF in peritoneal macrophages. Deletion of both the acidic loop and the β-hairpin completely abrogates IL-12 secretion. Insertion of the T. gondii acidic loop and β-hairpin into yeast profilin is sufficient to generate TLR11-dependent signaling. Substitution of the acidic loop in TgPRF with the homologous loop from the apicomplexan parasite Cryptosporidium parvum does not affect TLR11-dependent IL-12 secretion, while substitution with the acidic loop from Plasmodium falciparum results in reduced but significant IL-12 secretion. We conclude that the parasite-specific motif in TgPRF is the key molecular pattern recognized by TLR11. Unlike other profilins, TgPRF slows nucleotide exchange on monomeric rabbit actin and binds rabbit actin weakly. The putative TgPRF actin-binding surface includes the β-hairpin and diverges widely from the actin-binding surfaces of vertebrate profilins.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures


References
-
- Montoya JG, Liesenfeld O. Toxoplasmosis. Lancet. 2004;363:1965–1976. - PubMed
-
- Dobrowolski JM, Sibley LD. Toxoplasma invasion of mammalian cells is powered by the actin cytoskeleton of the parasite. Cell. 1996;84:933–939. - PubMed
-
- Schmitz S, Grainger M, Howell S, Calder LJ, Gaeb M, Pinder JC, Holder AA, Veigel C. Malaria parasite actin filaments are very short. J Mol Biol. 2005;349:113–125. - PubMed
-
- Schuler H, Mueller AK, Matuschewski K. Unusual properties of Plasmodium falciparum actin: new insights into microfilament dynamics of apicomplexan parasites. FEBS Lett. 2005;579:655–660. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
