

Review
doi: 10.1016/j.gde.2010.08.006.
Epub 2010 Sep 28.
Frequency control of cell cycle oscillators
Affiliations
- PMID: 20851595
- PMCID: PMC3522487
- DOI: 10.1016/j.gde.2010.08.006
Item in Clipboard
Review
Frequency control of cell cycle oscillators
Curr Opin Genet Dev.
2010 Dec.
Abstract
The cell cycle oscillator, based on a core negative feedback loop and modified extensively by positive feedback, cycles with a frequency that is regulated by environmental and developmental programs to encompass a wide range of cell cycle times. We discuss how positive feedback allows frequency tuning, how size and morphogenetic checkpoints regulate oscillator frequency, and how extrinsic oscillators such as the circadian clock gate cell cycle frequency. The master cell cycle regulatory oscillator in turn controls the frequency of peripheral oscillators controlling essential events. A recently proposed phase-locking model accounts for this coupling.
Copyright © 2010 Elsevier Ltd. All rights reserved.
Figures
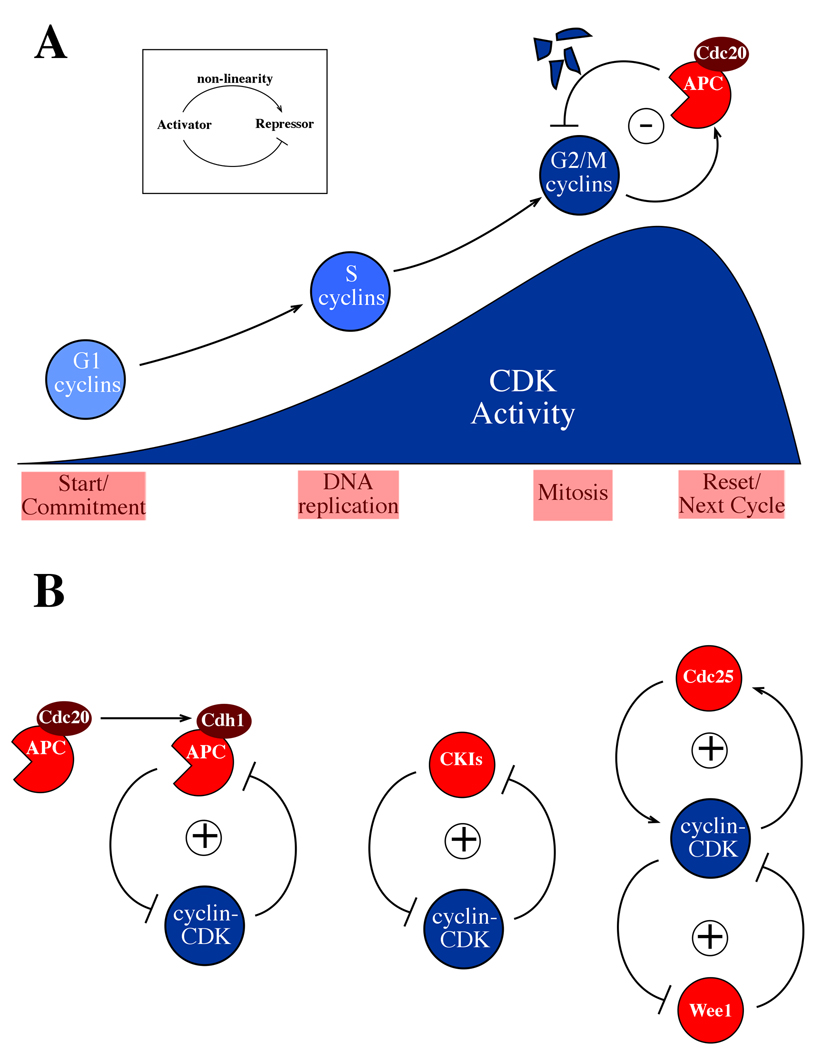
Positive and negative feedback loops in the cyclin-CDK oscillator. A Inset: a negative feedback loop which can give rise to oscillations. Such an oscillator is thought to form the core of eukaryotic cell cycles, with cyclin-Cyclin Dependent Kinase (cyclin-CDK) acting as activator, Anaphase Promoting Complex-Cdc20 (APC-Cdc20) acting as repressor, and non-linearity in APC-Cdc20 activation preventing the system from settling into a steady state. Below is shown the cyclin-CDK machinery in eukaryotic cell cycles. CDKs, present throughout the cell cycle, require the binding of a cyclin subunit for activity. These cyclin partners can also determine the localization of the complex and its specificity for targets. At the beginning of the cell cycle, cyclin-CDK activity is low, and ramps up over most of the cycle. Early cyclins trigger production of later cyclins and these later cyclins then turn off the earlier cyclins, so that control is passed from one set of cyclin-CDKs to the next. The last set of cyclins to be activated, the G2/M-phase cyclins, initiate mitosis, and also initiate their own destruction by activating the APC-Cdc20 negative feedback loop. APC-Cdc20 targets the G2/M-phase cyclins for destruction, resetting the cell to a low-CDK activity state, ready for the next cycle. B Positive feedback is added to the oscillator in multiple ways. Left: a highly-conserved but non-essential mechanism consists of “handoff” of cyclin proteolysis from APC-Cdc20 to APC-Cdh1. Cdh1 is a relative of Cdc20 which activates the APC late in mitosis and into the ensuing G1. Cdh1 is inhibited by cyclin-CDK activity, resulting in mutual inhibition (which is logically equivalent to positive feedback). Middle: antagonism between cyclin-CDK and stoichiometric CDK inhibitors (CKIs) results in positive feedback. These loops stabilize high- and low-CDK activity states. Right: a double positive feedback loop comprising CDK-mediated inhibition of the Wee1 kinase (which inhibits CDK) and activation of the Cdc25 phosphatase (which activates CDK by removing the phosphorylation added by Wee1) is proposed to stabilize intermediate CDK activity found in mid-cycle, and an alternative stable state of high mitotic CDK activity.
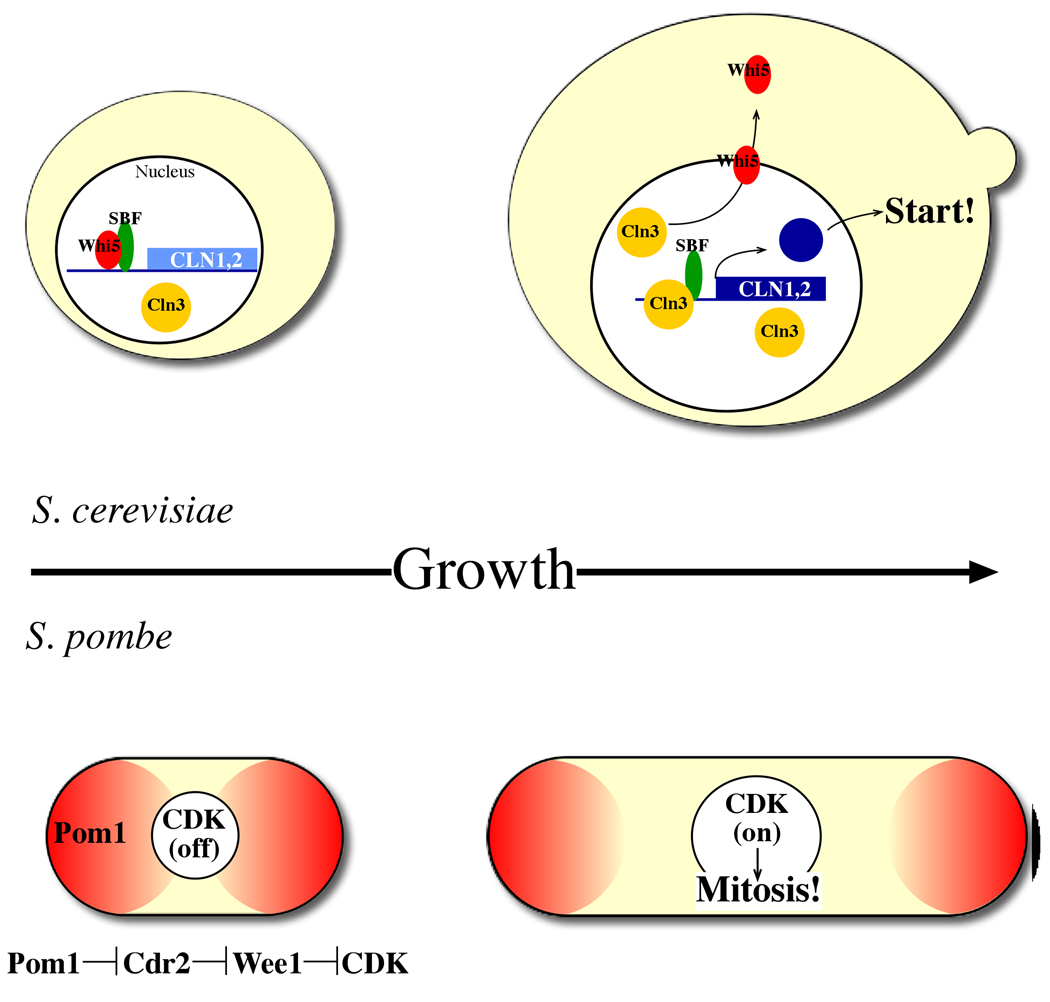
Size control mechanisms in budding and fission yeasts. Top: in S. cerevisiae, size control operates in G1. Transcription of many genes, including the G1/S cyclins (Cln1,2) is controlled by the SBF and MBF transcription factors (Swi4/Swi6 and Swi4/Mbp1, respectively). The Whi5 repressor inhibits this transcription until it is exported from the nucleus by the most upstream G1 cyclin, Cln3, in response to sufficient cell size. Cln3 thus relieves transcriptional inhibition, promoting Cln1,2 expression and subsequent cell cycle Start. Actual size “measurement” was recently proposed to operate through direct binding of Cln3 to the SCB target sequences of Swi4/Swi6, with Start occurring upon titration of these sites by Cln3. Bottom: in S. pombe, size control operates in G2. Pom1, localized to cell poles, indirectly inhibits CDK activity (through inhibition of Cdr2, which inhibits Wee1, which in turn inhibits CDK). As the cell elongates, the concentration of Pom1 at the center of the cell (where the nucleus is located) drops, allowing CDK activation leading to mitosis.
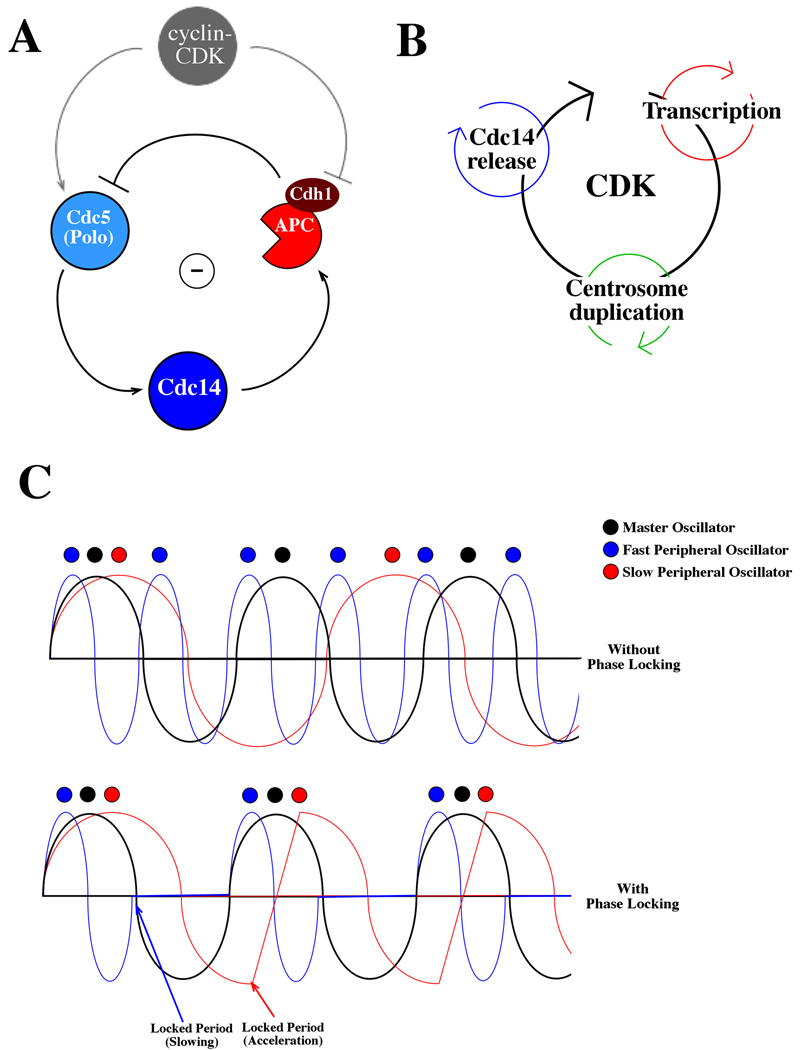
A phase-locking model for entrainment of peripheral oscillators to the cyclin-CDK ocillator. A Molecular mechanism of the Cdc14 release oscillator. The mitotic phosphatase Cdc14 is activated upon release from sequestration in the nucleolus. This release is controlled by a negative feedback loop in which Cdc14 release, promoted by the polo kinase Cdc5, activates APC-Cdh1, which then promotes Cdc5 degradation, allowing Cdc14 resequestration. This negative feedback oscillator is entrained to the cyclin-CDK cycle at multiple points: by cyclin-CDK promotion of CDC5 transcription and Cdc5 kinase activation, and by cyclin-CDK inhibition of Cdh1 activity. B Schematic of multiple peripheral oscillators coupled to the CDK oscillator in budding yeast. As described above, coupling entrains such peripheral oscillators to cell cycle progression; peripheral oscillators also feed back on the cyclin-CDK oscillator itself. For example, major genes in the periodic transcription program include most cyclins, CDC20, and CDC5; Cdc14 directly promotes establishment of the low-cyclin-CDK positive feedback loop by activating Cdh1 and Sic1 as well as more indirectly antagonizing cyclin-CDK activity by dephosphorylating cyclin-CDK targets; the centrosome and budding cycles could communicate with the cyclin-CDK cycle via the spindle integrity and morphogenesis checkpoints. C Oscillator coupling ensures once-per-cell-cycle occurrence of events. Three hypothetical oscillators are shown: a master cycle in black, a faster peripheral cycle in blue, and a slower peripheral cycle in red. In the absence of phase-locking (top), the oscillators trigger events (colored circles) without a coherent phase relationship. In the presence of oscillator coupling (bottom), the peripheral oscillators are slowed or accelerated within their critical periods to produce a locked phase relationship, with events occurring once and only once within each master cycle.
Similar articles
-
Circadian clocks and cell division: what's the pacemaker?Cell Cycle. 2010 Oct 1;9(19):3864-73. doi: 10.4161/cc.9.19.13205. Epub 2010 Oct 1. Cell Cycle. 2010. PMID: 20890114 Free PMC article.
-
Contribution of membrane-associated oscillators to biological timing at different timescales.Front Physiol. 2024 Jan 9;14:1243455. doi: 10.3389/fphys.2023.1243455. eCollection 2023. Front Physiol. 2024. PMID: 38264332 Free PMC article.
-
A detailed map of coupled circadian clock and cell cycle with qualitative dynamics validation.BMC Bioinformatics. 2021 May 11;22(1):240. doi: 10.1186/s12859-021-04158-9. BMC Bioinformatics. 2021. PMID: 33975535 Free PMC article.
-
Regulation of tissue regeneration by the circadian clock.Eur J Neurosci. 2021 Jun;53(11):3576-3597. doi: 10.1111/ejn.15244. Epub 2021 May 14. Eur J Neurosci. 2021. PMID: 33893679 Review.
-
Biological timing and the clock metaphor: oscillatory and hourglass mechanisms.Chronobiol Int. 2001 May;18(3):329-69. doi: 10.1081/cbi-100103961. Chronobiol Int. 2001. PMID: 11475408 Review.
Cited by
-
The Inner Nuclear Membrane Protein Src1 Is Required for Stable Post-Mitotic Progression into G1 in Aspergillus nidulans.PLoS One. 2015 Jul 6;10(7):e0132489. doi: 10.1371/journal.pone.0132489. eCollection 2015. PLoS One. 2015. PMID: 26147902 Free PMC article.
-
Roles for the Histone Modifying and Exchange Complex NuA4 in Cell Cycle Progression in Drosophila melanogaster.Genetics. 2016 Jul;203(3):1265-81. doi: 10.1534/genetics.116.188581. Epub 2016 May 16. Genetics. 2016. PMID: 27184390 Free PMC article.
-
Oscillation of clock and clock controlled genes induced by serum shock in human breast epithelial and breast cancer cells: regulation by melatonin.Breast Cancer (Auckl). 2012;6:137-50. doi: 10.4137/BCBCR.S9673. Epub 2012 Sep 13. Breast Cancer (Auckl). 2012. PMID: 23012497 Free PMC article.
-
Hippo signaling is intrinsically regulated during cell cycle progression by APC/CCdh1.Proc Natl Acad Sci U S A. 2019 May 7;116(19):9423-9432. doi: 10.1073/pnas.1821370116. Epub 2019 Apr 18. Proc Natl Acad Sci U S A. 2019. PMID: 31000600 Free PMC article.
-
Design of Oscillatory Networks through Post-Translational Control of Network Components.Synth Biol Eng. 2023 Jun;1(1):10004. doi: 10.35534/sbe.2023.10004. Epub 2023 Mar 13. Synth Biol Eng. 2023. PMID: 38590452 Free PMC article.
References
-
- Tyson JJ, Chen KC, Novak B. Sniffers, buzzers, toggles and blinkers: dynamics of regulatory and signaling pathways in the cell. Curr Opin Cell Biol. 2003;15:221–231. - PubMed
-
- Felix MA, Labbe JC, Doree M, Hunt T, Karsenti E. Triggering of cyclin degradation in interphase extracts of amphibian eggs by cdc2 kinase. Nature. 1990;346:379–382. - PubMed
-
- Dunlap JC, Loros JJ, Colot HV, Mehra A, Belden WJ, Shi M, Hong CI, Larrondo LF, Baker CL, Chen CH, et al. A circadian clock in Neurospora: how genes and proteins cooperate to produce a sustained, entrainable, and compensated biological oscillator with a period of about a day. Cold Spring Harb Symp Quant Biol. 2007;72:57–68. - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
