

A bird brain's view of auditory processing and perception
- PMID: 20851756
- PMCID: PMC3065528
- DOI: 10.1016/j.heares.2010.08.008
A bird brain's view of auditory processing and perception
Abstract
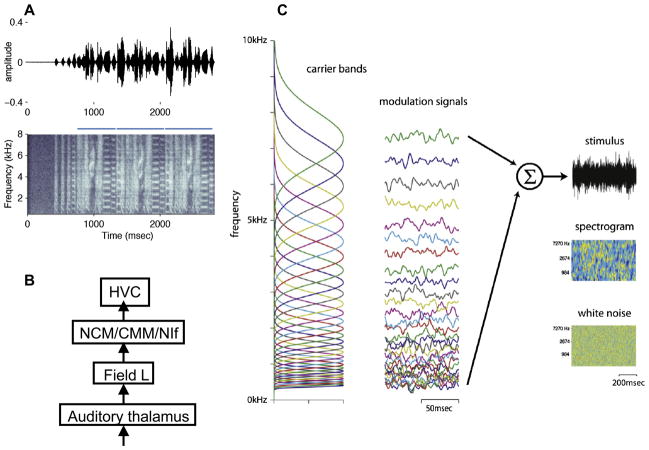
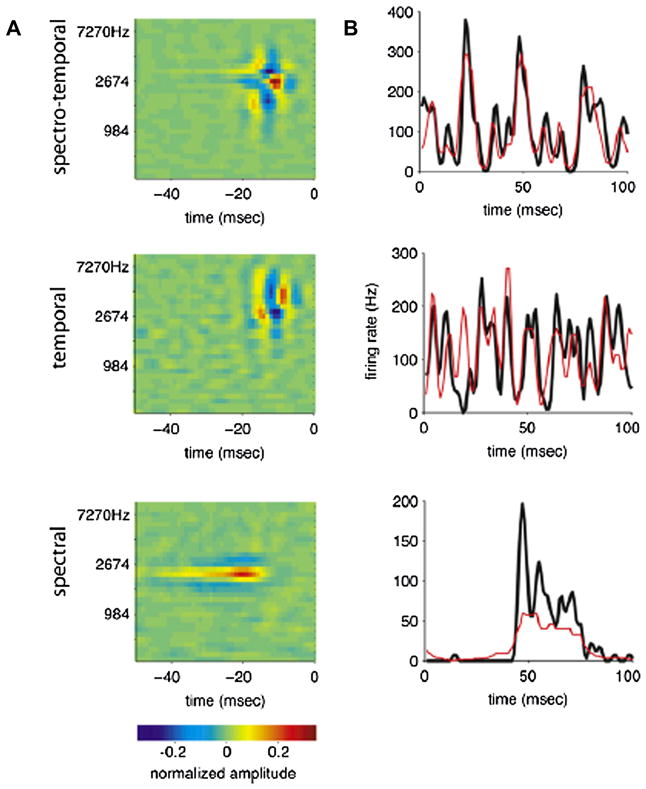
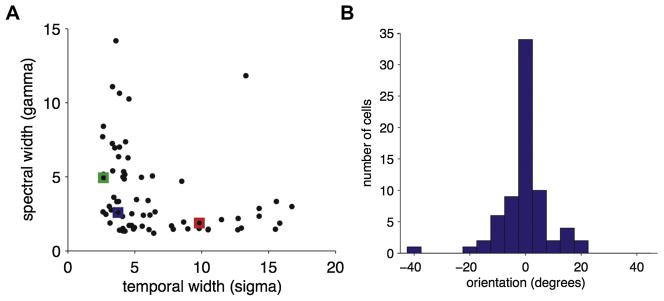
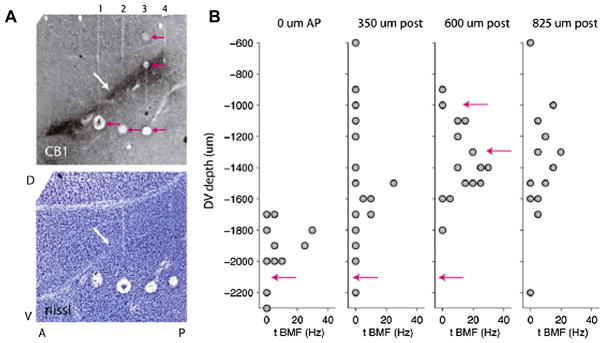
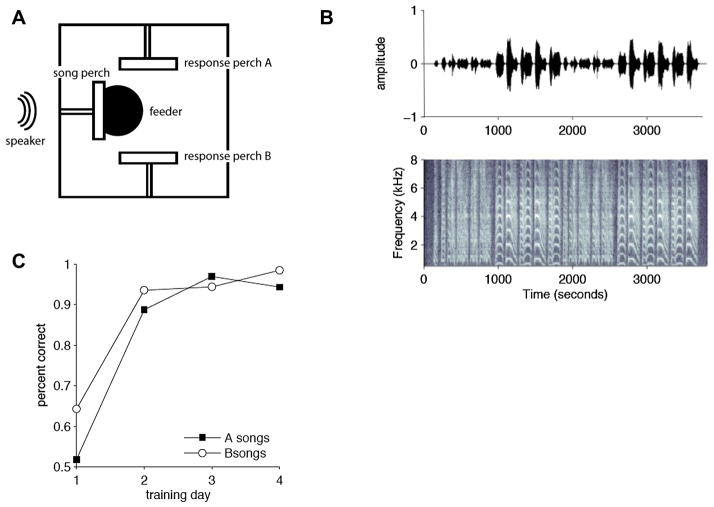
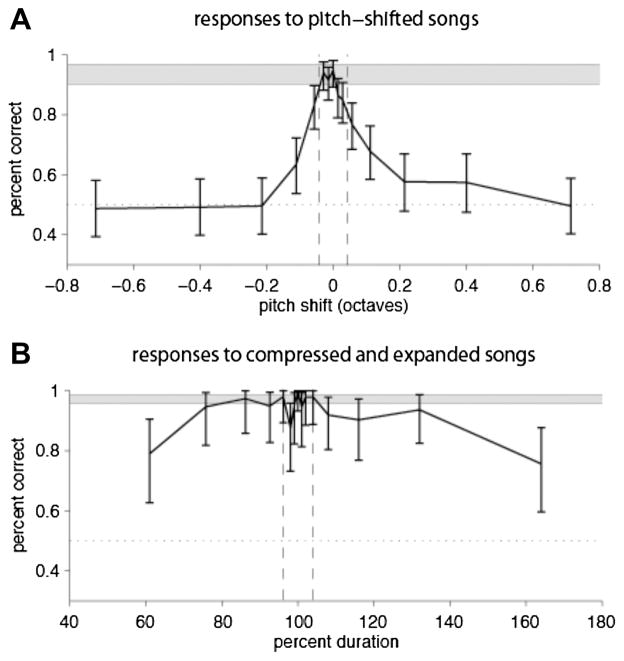
By studying the primary forebrain auditory area of songbirds, field L, using a song-inspired synthetic stimulus and reverse correlation techniques, we found a surprisingly systematic organization of this area, with nearly all neurons narrowly tuned along the spectral dimension, the temporal dimension, or both; there were virtually no strongly orientation-sensitive cells, and in the areas that we recorded, cells broadly tuned in both time and frequency were rare. In addition, cells responsive to fast temporal frequencies predominated only in the field L input layer, suggesting that neurons with fast and slow responses are concentrated in different regions. Together with other songbird data and work from chicks and mammals, these findings suggest that sampling a range of temporal and spectral modulations, rather than orientation in time-frequency space, is the organizing principle of forebrain auditory sensitivity. We then examined the role of these acoustic parameters important to field L organization in a behavioral task. Birds' categorization of songs fell off rapidly when songs were altered in frequency, but, despite the temporal sensitivity of field L neurons, the same birds generalized well to songs that were significantly changed in timing. These behavioral data point out that we cannot assume that animals use the information present in particular neurons without specifically testing perception.
Copyright © 2010 Elsevier B.V. All rights reserved.
Figures
References
-
- Amin N, Theunissen FE. Selectivity for natural sounds in the songbird auditory forebrain is strongly shaped by the acoustic environment. Society for Neuroscience Abstract. 2008:99.8.
-
- Avedaño C, Deng L, Hermansky H, Gold B. Speech Processing in the Auditory System. New York, NY: Springer-Verlag; 2004. The analysis and representation of speech.
-
- Brenowitz EA. Altered perception of species-specific song by female birds after lesions of a forebrain nucleus. Science. 1991;251(4991):303–5. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
