

Review
doi: 10.1007/s12015-010-9191-9.
Functional characterization of stem cell activity in the mouse mammary gland
Affiliations
- PMID: 20853073
- PMCID: PMC3492888
- DOI: 10.1007/s12015-010-9191-9
Item in Clipboard
Review
Functional characterization of stem cell activity in the mouse mammary gland
Stem Cell Rev Rep.
2011 Jun.
Abstract
Any portion of the mouse mammary gland is capable of recapitulating a clonally derived complete and functional mammary tree upon transplantation into an epithelial divested mammary fat-pad of a recipient host. As such, it is an ideal model tissue for the study somatic stem cell function. This review will outline what is known regarding the function of stem/progenitor cells in the mouse mammary gland, including how progenitor populations can be functionally defined, the evidence for and potential role of selective DNA strand segregation, and the role of the niche in maintaining and controlling stem cell function.
Conflict of interest statement
Figures
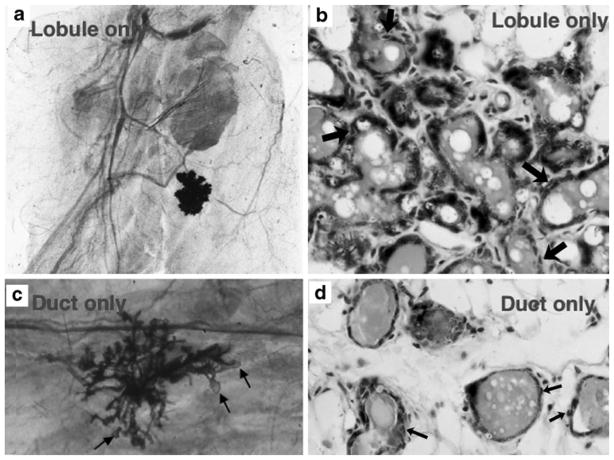
a and b Example of a lobule-limited outgrowth. a Whole mounted mammary fat pad of lobule-limited outgrowth in a full term pregnant host developed from a limiting dilution inoculum of mammary cells. b A cross-section of a lobule limited outgrowth reveals presence of luminal and myoepithelial cells (arrows). c and d Example of a duct-limited outgrowth. c A whole-mounted mammary fat pad of a duct limited outgrowth from a full term pregnant host with attendant terminal end buds (arrows) and no secretory lobule development. d Section through the duct-limited growth shown in (c) demonstrating the presence of both luminal and myoepithelial cells (arrows)
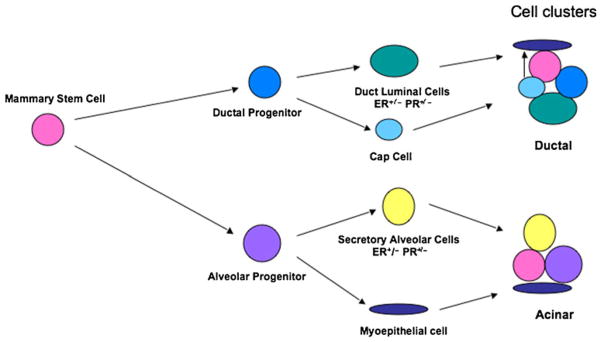
Model of stem/progenitor cellular hierarchy in the mouse mammary gland based on in-vivo transplantation. The pluripotent stem cell gives rise to duct-limited and lobule-limited progenitors. Both progenitor populations are multipotent. The lobule progenitor gives rise to ER+/− and PR+/− luminal epithelial cells and myeopeithelial cells. Duct-limited progenitors also give rise to ER+/− and PR+/− luminal epithelial cells as well as the cap cells during ductal elongation. The cap cells ultimately differentiate into the myoepithelial cells of the ducts. Importantly, stem, lobule-limited, and duct-limited functions all seem to be required for complete mammary development
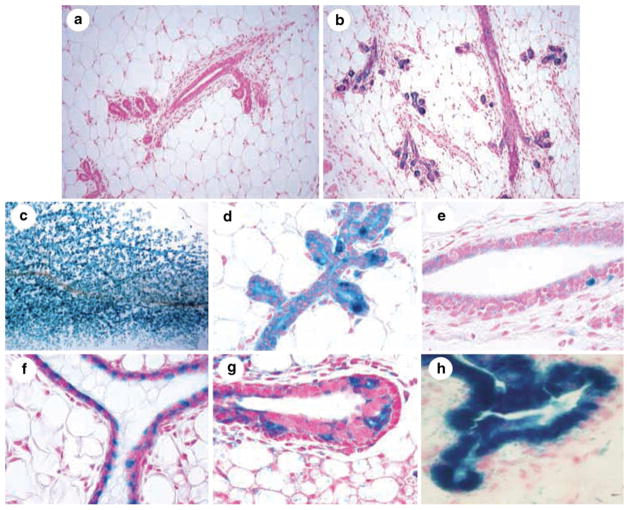
Identification of PI-MEC in WC/R26-lacZ mice. a Tissue section of a nulliparous WC/R26-lacZ mouse is negative for lacZ expression. b Tissue sections from parous WC/R26-lacZ mouse following involution demonstrates presence of lacZ positive cells (i. e. PI-MEC) near the extremities of ducts. c–e Mammary whole mount (c) and tissue sections (d and e) of WC/R26-lacZ mice at day 8 of second gestation period reveal PI-MEC proliferate and function as alveolar progenitors (c and d) in subsequent pregnancies but remain largely absent and inactive in primary ducts (e). f–h Transplantation of PI-MEC results in second generation outgrowths that contain lacZ positive cells along primary ducts (f) and in the terminal end bud (g). h PI-MEC continue to function as alveolar progenitors in second generation outgrowths and contribute blue progeny to developing secretory acini
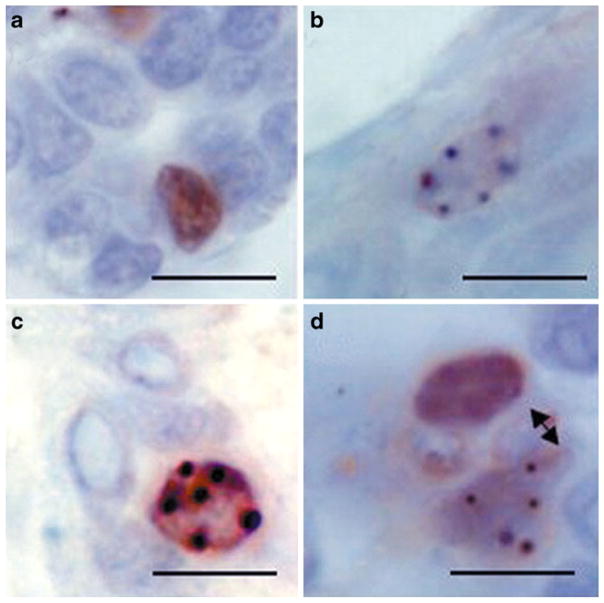
Label retaining epithelial cells in the mouse mammary gland asymmetrically divide their DNA and retain their template strand. Mice were treated with 3HTdR on 5 consecutive days 10 days following transplantation of mammary fragments into their epithelium divested #4 and #9 mammary glands. Following a 5 week chase period, mice were treated with BrdU on 2 consecutive days. a and b Cells that are single positive for either BrdU (a) or 3HTdR (b). c A cell that is double positive for BrdU and 3HTdR. d A juxtaposed single positive BrdU cell and a 3HTdR positive cell possibly resulting from a recent asymmetric division. Scale bar=10 μM
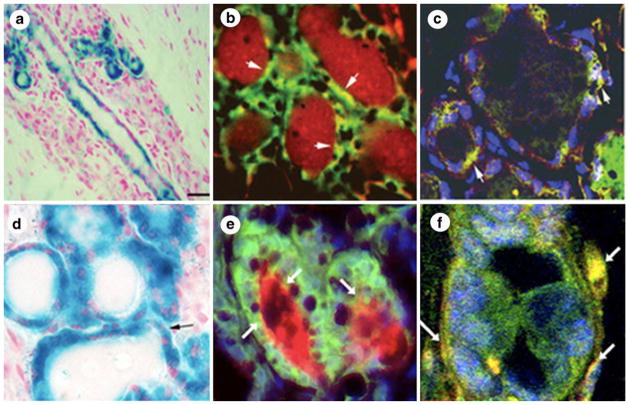
Testicular cells (a–c) and NSC (d–f) from WC/R26-lacZ mice are reprogrammed by the mammary microenvironment to adopt a mammary epithelial differentiation repertoire. Testicular cells or NSC from WC/R26-lacZ mice were mixed in a 1:1 ratio with wild-type FVB mammary epithelial cells. a and d Tissue sections reveal lacZ+ cells within mammary structures. b and e Co-localization of beta-gal (green) and caseins (red) demonstrates testicular (b) and NSC (e) produce secretory luminal epithelial cell progeny. c and f Co-localization of beta-gal (green) and smooth muscle actin (red) demonstrates testicular (c) and NSC (f) produce myoepithelial progeny
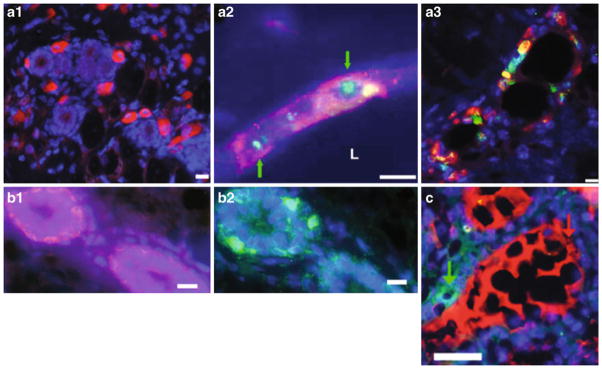
When mixed with normal mouse mammary epthilelial cells (MEC), human embryonal carcinoma (NT2)-derived cells are incorporated into the mammary epithelium and differentiate. a1 CD133-positive NT2 cells (red) are present within the confines of the fat pad containing regenerated mammary ducts. a2 human NT2 cells are present within the mammary outgrowths as determined by human-specific FISH (green, nuclear; identified with green arrows) and human-specific immunocytochemical staining for CD133 (red). a3 NT2 cells in regenerated mammary epithelium, as determined by expression of human CD133 (red), differentiate into luminal epithelial cells and express ER-α (green). b1 and b2 the location of basal cells (b1, red) expressing hK14 and mouse K14 (b2, green) in consecutive sections showing the same chimeric duct. c human and mouse proteins are secreted into the same lumen. Simultaneous staining of chimeric mammary outgrowth for human α lactalbumin (green) and mouse caseins (red) shows that production and secretion of both mouse and human milk proteins are present in the same chimeric duct. All fluorescent sections are counterstained with DAPI (blue). Scale bars, 20μm (a1), 15 μm (a2 and a3), 10 μm (b1 and b2), and 25 μm (c)
References
-
- Deome K, Faulkin LJ, Bern H, Blair P. Development of mammary tumors from hyperplastic alveolar nodules transplanted into gland-free mammary fat pads of female C3H mice. Cancer Research. 1959;19:515–520. - PubMed
-
- Faulkin LJ, Jr, Deome KB. Regulation of growth and spacing of gland elements in the mammary fat pad of the C3H mouse. Journal of the National Cancer Institute. 1960;24:953–969. - PubMed
-
- Daniel CW, Deome KB. Growth of mouse mammary glands in vivo after monolayer culture. Science. 1965;149:634–636. - PubMed
-
- Daniel CW, Aidells BD, Medina D, Faulkin LJ., Jr Unlimited division potential of precancerous mouse mammary cells after spontaneous or carcinogen-induced transformation. Federation Proceedings. 1975;34:64–67. - PubMed
-
- Smith GH, Medina D. A morphologically distinct candidate for an epithelial stem cell in mouse mammary gland. Journal of Cell Science. 1988;90(Pt 1):173–183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
