

Cooperative action of multiple cis-acting elements is required for N-myc expression in branchial arches: specific contribution of GATA3
- PMID: 20855530
- PMCID: PMC2976382
- DOI: 10.1128/MCB.00353-09
Cooperative action of multiple cis-acting elements is required for N-myc expression in branchial arches: specific contribution of GATA3
Erratum in
- Mol Cell Biol. 2012 Dec;32(24):5152-3
Abstract
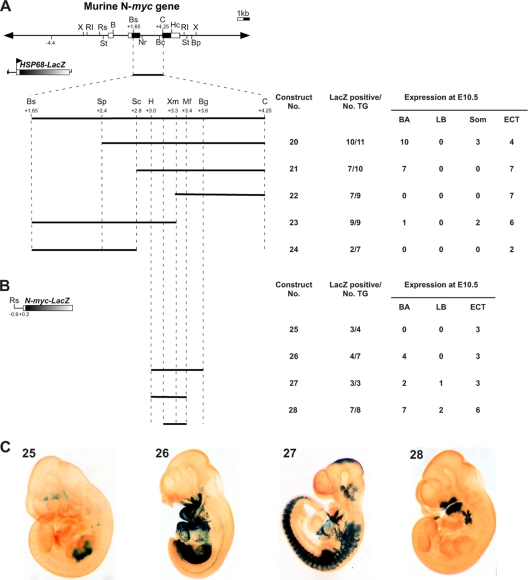
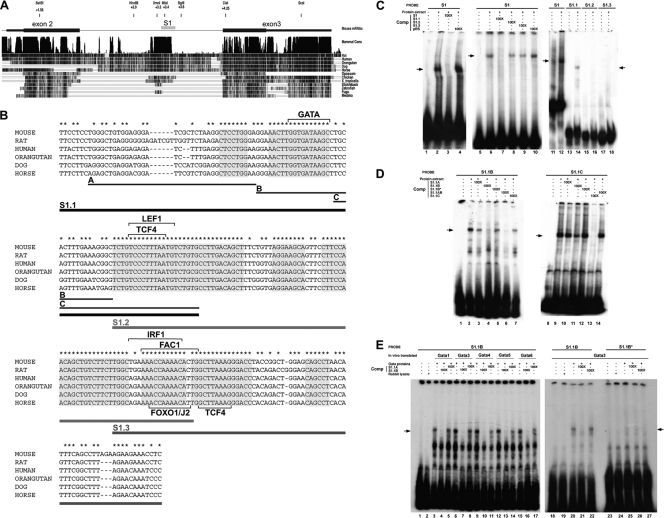
The precise expression of the N-myc proto-oncogene is essential for normal mammalian development, whereas altered N-myc gene regulation is known to be a determinant factor in tumor formation. Using transgenic mouse embryos, we show that N-myc sequences from kb -8.7 to kb +7.2 are sufficient to reproduce the N-myc embryonic expression profile in developing branchial arches and limb buds. These sequences encompass several regulatory elements dispersed throughout the N-myc locus, including an upstream limb bud enhancer, a downstream somite enhancer, a branchial arch enhancer in the second intron, and a negative regulatory element in the first intron. N-myc expression in the limb buds is under the dominant control of the limb bud enhancer. The expression in the branchial arches necessitates the interplay of three regulatory domains. The branchial arch enhancer cooperates with the somite enhancer region to prevent an inhibitory activity contained in the first intron. The characterization of the branchial arch enhancer has revealed a specific role of the transcription factor GATA3 in the regulation of N-myc expression. Together, these data demonstrate that correct N-myc developmental expression is achieved via cooperation of multiple positive and negative regulatory elements.
Figures








References
-
- Alt, F. W., R. DePinho, K. Zimmerman, E. Legouy, K. Hatton, P. Ferrier, A. Tesfaye, G. Yancopoulos, and P. Nisen. 1986. The human myc gene family. Cold Spring Harbor Symp. Quant. Biol. 51:931-941. - PubMed
-
- Aubry, S., and J. Charron. 2000. N-Myc shares cellular functions wih c-Myc. DNA Cell Biol. 19:353-364. - PubMed
-
- Baena, E., M. Ortiz, A. C. Martinez, and I. M. de Alboran. 2007. c-Myc is essential for hematopoietic stem cell differentiation and regulates Lin(-)Sca-1(+)c-Kit(-) cell generation through p21. Exp. Hematol. 35:1333-1343. - PubMed
-
- Ball, N. S., and P. H. King. 1997. Neuron-specific Hel-N1 and HuD as novel molecular markers of neuroblastoma: a correlation of HuD messenger RNA levels with favorable prognostic features. Clin. Cancer Res. 3:1859-1865. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases