

Loss of STOP protein impairs peripheral olfactory neurogenesis
- PMID: 20856814
- PMCID: PMC2939889
- DOI: 10.1371/journal.pone.0012753
Loss of STOP protein impairs peripheral olfactory neurogenesis
Abstract
Background: STOP (Stable Tubulin-Only Polypeptide) null mice show behavioral deficits, impaired synaptic plasticity, decrease in synaptic vesicular pools and disturbances in dopaminergic transmission, and are considered a neurodevelopmental model of schizophrenia. Olfactory neurons highly express STOP protein and are continually generated throughout life. Experimentally-induced loss of olfactory neurons leads to epithelial regeneration within two months, providing a useful model to evaluate the role played by STOP protein in adult olfactory neurogenesis.
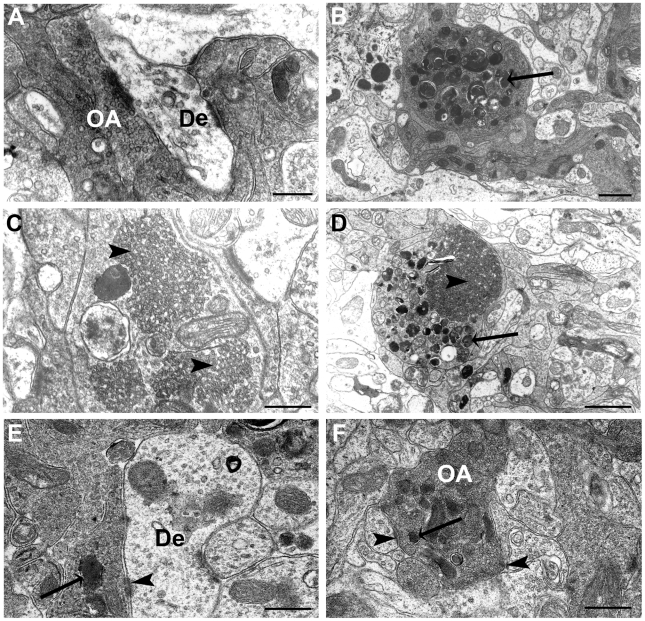
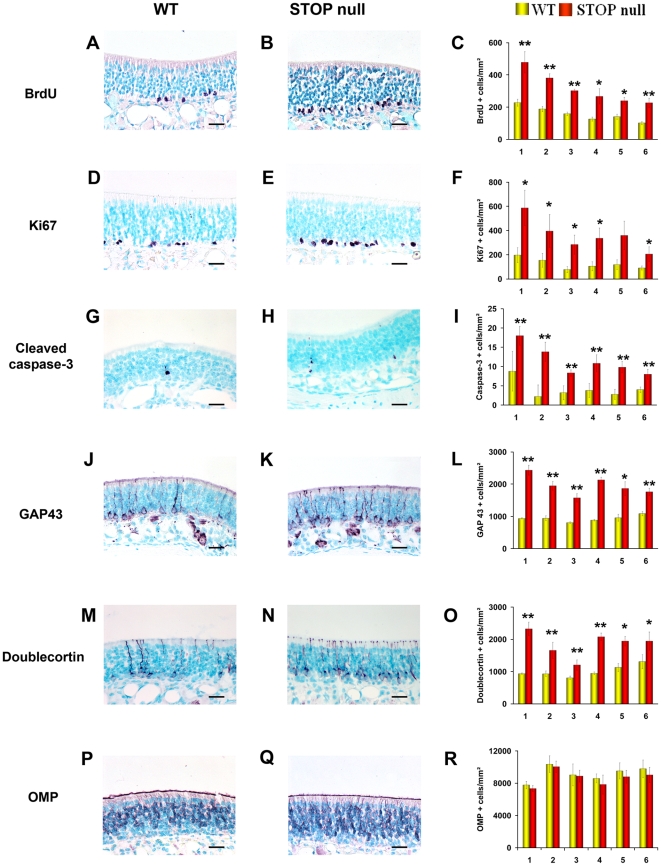
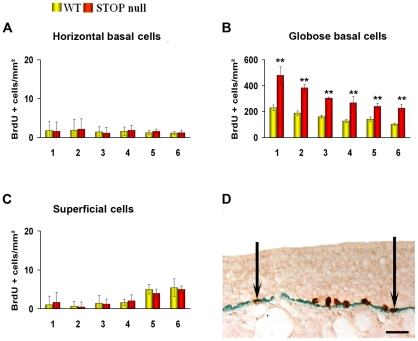
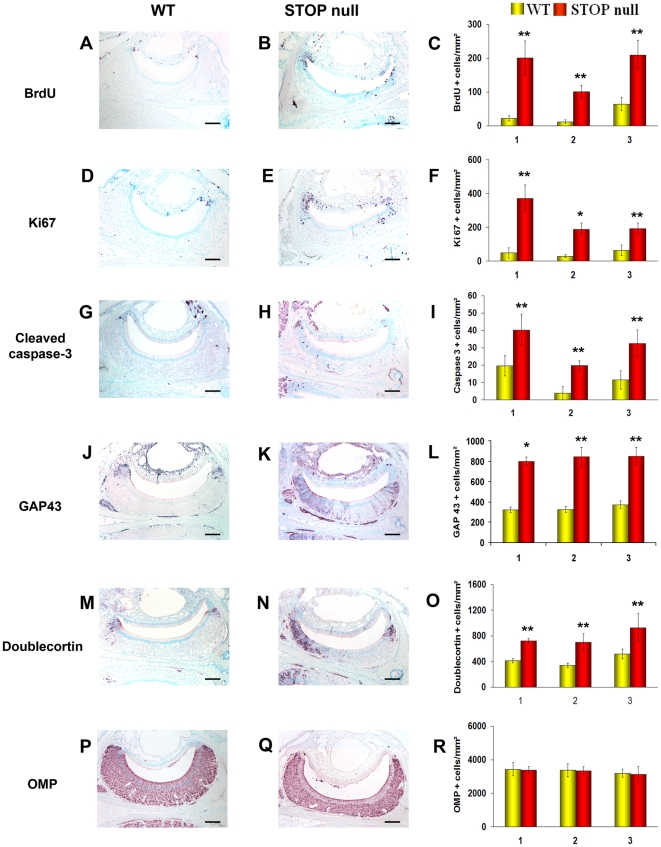
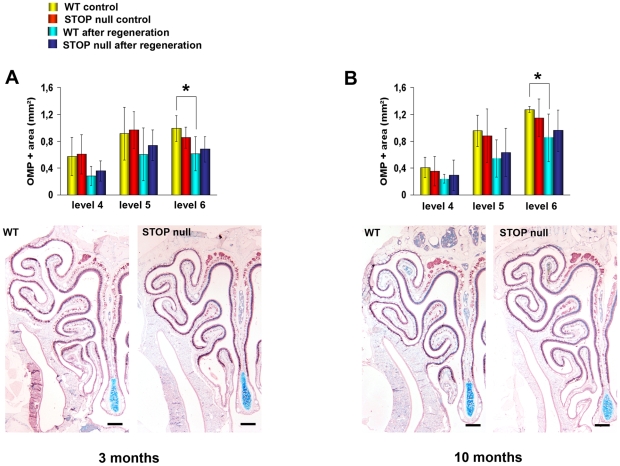
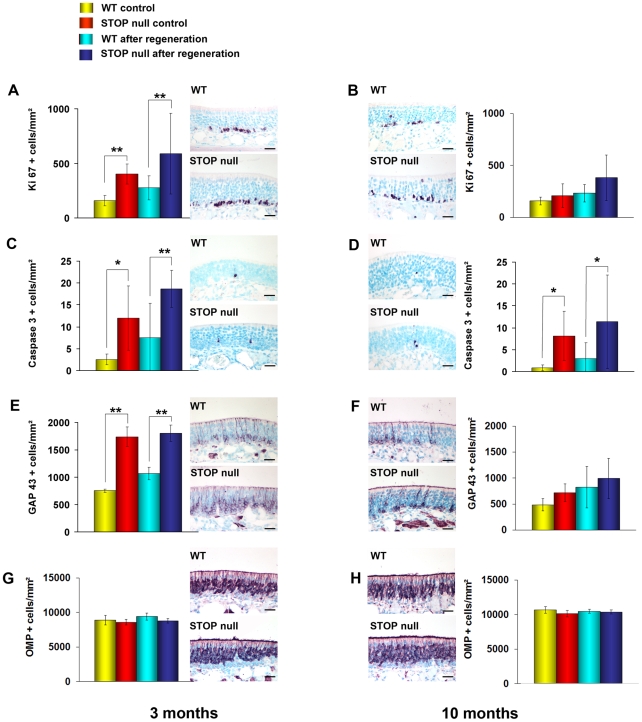
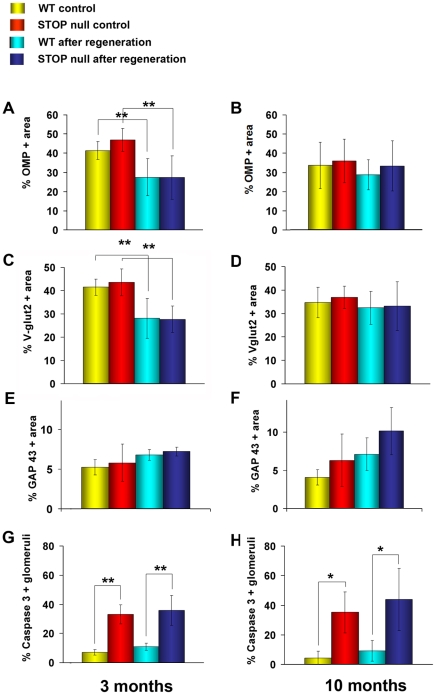
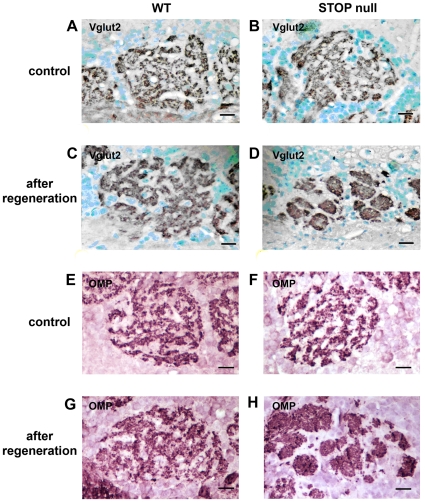
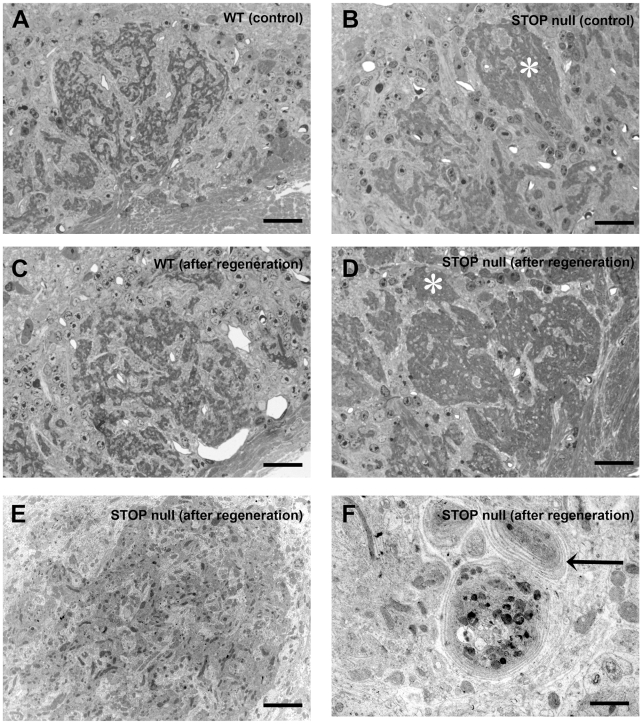
Methodology/principal findings: Immunocytochemistry and electron microscopy were used to study the structure of the glomerulus in the main olfactory bulb and neurogenesis in the neurosensorial epithelia. In STOP null mice, olfactory neurons showed presynaptic swellings with tubulovesicular profiles and autophagic-like structures. In olfactory and vomeronasal epithelia, there was an increase in neurons turnover, as shown by the increase in number of proliferating, apoptotic and immature cells with no changes in the number of mature neurons. Similar alterations in peripheral olfactory neurogenesis have been previously described in schizophrenia patients. In STOP null mice, regeneration of the olfactory epithelium did not modify these anomalies; moreover, regeneration resulted in abnormal organisation of olfactory terminals within the olfactory glomeruli in STOP null mice.
Conclusions/significance: In conclusion, STOP protein seems to be involved in the establishment of synapses in the olfactory glomerulus. Our results indicate that the olfactory system of STOP null mice is a well-suited experimental model (1) for the study of the mechanism of action of STOP protein in synaptic function/plasticity and (2) for pathophysiological studies of the mechanisms of altered neuronal connections in schizophrenia.
Conflict of interest statement
Figures
Similar articles
-
Characterization of olfactory bulb glomeruli in schizophrenia.Schizophr Res. 2005 Sep 15;77(2-3):229-39. doi: 10.1016/j.schres.2005.04.022. Schizophr Res. 2005. PMID: 15946825
-
Altered expression of synaptic protein mRNAs in STOP (MAP6) mutant mice.J Psychopharmacol. 2007 Aug;21(6):635-44. doi: 10.1177/0269881106068825. Epub 2006 Oct 18. J Psychopharmacol. 2007. PMID: 17050659
-
CX₃CR1 deficiency exacerbates neuronal loss and impairs early regenerative responses in the target-ablated olfactory epithelium.Mol Cell Neurosci. 2011 Nov;48(3):236-45. doi: 10.1016/j.mcn.2011.08.004. Epub 2011 Aug 16. Mol Cell Neurosci. 2011. PMID: 21871566
-
Development of the mammalian main olfactory bulb.Development. 2022 Feb 1;149(3):dev200210. doi: 10.1242/dev.200210. Epub 2022 Feb 11. Development. 2022. PMID: 35147186 Free PMC article. Review.
-
A lifetime of neurogenesis in the olfactory system.Front Neurosci. 2014 Jun 26;8:182. doi: 10.3389/fnins.2014.00182. eCollection 2014. Front Neurosci. 2014. PMID: 25018692 Free PMC article. Review.
Cited by
-
Structural basis for the association of MAP6 protein with microtubules and its regulation by calmodulin.J Biol Chem. 2013 Aug 23;288(34):24910-22. doi: 10.1074/jbc.M113.457267. Epub 2013 Jul 6. J Biol Chem. 2013. PMID: 23831686 Free PMC article.
-
Altered circadian activity and sleep/wake rhythms in the stable tubule only polypeptide (STOP) null mouse model of schizophrenia.Sleep. 2021 Apr 9;44(4):zsaa237. doi: 10.1093/sleep/zsaa237. Sleep. 2021. PMID: 33186470 Free PMC article.
-
Changes in ADAR RNA editing patterns in CMV and ZIKV congenital infections.BMC Genomics. 2023 Nov 15;24(1):685. doi: 10.1186/s12864-023-09778-4. BMC Genomics. 2023. PMID: 37968596 Free PMC article.
-
Beyond Neuronal Microtubule Stabilization: MAP6 and CRMPS, Two Converging Stories.Front Mol Neurosci. 2021 May 5;14:665693. doi: 10.3389/fnmol.2021.665693. eCollection 2021. Front Mol Neurosci. 2021. PMID: 34025352 Free PMC article.
-
Microtubule-associated protein 6 mediates neuronal connectivity through Semaphorin 3E-dependent signalling for axonal growth.Nat Commun. 2015 Jun 3;6:7246. doi: 10.1038/ncomms8246. Nat Commun. 2015. PMID: 26037503 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
