

Gingival fibroblasts as a promising source of induced pluripotent stem cells
- PMID: 20856871
- PMCID: PMC2939066
- DOI: 10.1371/journal.pone.0012743
Gingival fibroblasts as a promising source of induced pluripotent stem cells
Abstract
Background: Induced pluripotent stem (iPS) cells efficiently generated from accessible tissues have the potential for clinical applications. Oral gingiva, which is often resected during general dental treatments and treated as biomedical waste, is an easily obtainable tissue, and cells can be isolated from patients with minimal discomfort.
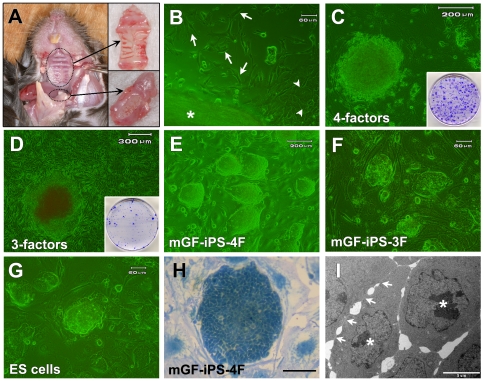
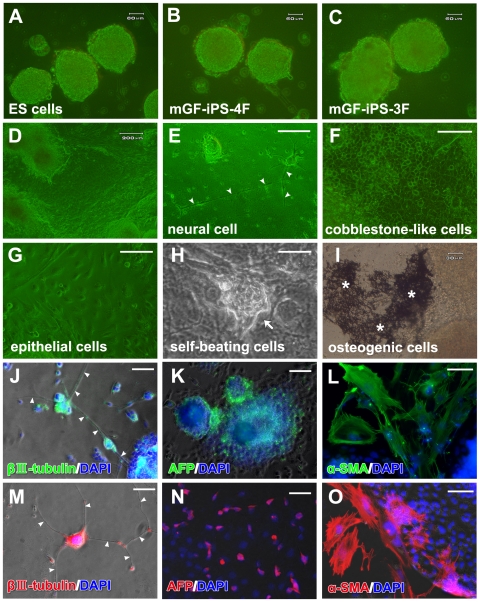
Methodology/principal findings: We herein demonstrate iPS cell generation from adult wild-type mouse gingival fibroblasts (GFs) via introduction of four factors (Oct3/4, Sox2, Klf4 and c-Myc; GF-iPS-4F cells) or three factors (the same as GF-iPS-4F cells, but without the c-Myc oncogene; GF-iPS-3F cells) without drug selection. iPS cells were also generated from primary human gingival fibroblasts via four-factor transduction. These cells exhibited the morphology and growth properties of embryonic stem (ES) cells and expressed ES cell marker genes, with a decreased CpG methylation ratio in promoter regions of Nanog and Oct3/4. Additionally, teratoma formation assays showed ES cell-like derivation of cells and tissues representative of all three germ layers. In comparison to mouse GF-iPS-4F cells, GF-iPS-3F cells showed consistently more ES cell-like characteristics in terms of DNA methylation status and gene expression, although the reprogramming process was substantially delayed and the overall efficiency was also reduced. When transplanted into blastocysts, GF-iPS-3F cells gave rise to chimeras and contributed to the development of the germline. Notably, the four-factor reprogramming efficiency of mouse GFs was more than 7-fold higher than that of fibroblasts from tail-tips, possibly because of their high proliferative capacity.
Conclusions/significance: These results suggest that GFs from the easily obtainable gingival tissues can be readily reprogrammed into iPS cells, thus making them a promising cell source for investigating the basis of cellular reprogramming and pluripotency for future clinical applications. In addition, high-quality iPS cells were generated from mouse GFs without Myc transduction or a specific system for reprogrammed cell selection.
Conflict of interest statement
Figures





Similar articles
-
The timing of retroviral silencing correlates with the quality of induced pluripotent stem cell lines.Biochim Biophys Acta. 2011 Feb;1810(2):226-35. doi: 10.1016/j.bbagen.2010.10.004. Epub 2010 Oct 20. Biochim Biophys Acta. 2011. PMID: 20965232
-
Induced pluripotent stem cell lines derived from human gingival fibroblasts and periodontal ligament fibroblasts.J Periodontal Res. 2011 Aug;46(4):438-47. doi: 10.1111/j.1600-0765.2011.01358.x. Epub 2011 Mar 29. J Periodontal Res. 2011. PMID: 21443752
-
Generation of mouse induced pluripotent stem cells from different genetic backgrounds using Sleeping beauty transposon mediated gene transfer.Exp Cell Res. 2012 Nov 15;318(19):2482-9. doi: 10.1016/j.yexcr.2012.07.014. Epub 2012 Jul 28. Exp Cell Res. 2012. PMID: 22846649
-
Emerging methods for preparing iPS cells.Jpn J Clin Oncol. 2012 Sep;42(9):773-9. doi: 10.1093/jjco/hys108. Epub 2012 Jul 23. Jpn J Clin Oncol. 2012. PMID: 22826352 Review.
-
iPS cells: a source of cardiac regeneration.J Mol Cell Cardiol. 2011 Feb;50(2):327-32. doi: 10.1016/j.yjmcc.2010.10.026. Epub 2010 Oct 30. J Mol Cell Cardiol. 2011. PMID: 21040726 Review.
Cited by
-
Intermittent compressive force induces cell cycling and reduces apoptosis in embryoid bodies of mouse induced pluripotent stem cells.Int J Oral Sci. 2022 Jan 4;14(1):1. doi: 10.1038/s41368-021-00151-3. Int J Oral Sci. 2022. PMID: 34980892 Free PMC article.
-
Human fetal dermal fibroblast-myeloid cell diversity is characterized by dominance of pro-healing Annexin1-FPR1 signaling.iScience. 2023 Aug 2;26(9):107533. doi: 10.1016/j.isci.2023.107533. eCollection 2023 Sep 15. iScience. 2023. PMID: 37636079 Free PMC article.
-
Stem Cells and Their Derivatives-Implications for Alveolar Bone Regeneration: A Comprehensive Review.Int J Mol Sci. 2021 Oct 29;22(21):11746. doi: 10.3390/ijms222111746. Int J Mol Sci. 2021. PMID: 34769175 Free PMC article. Review.
-
Tissue-specific promoter-based reporter system for monitoring cell differentiation from iPSCs to cardiomyocytes.Sci Rep. 2020 Feb 5;10(1):1895. doi: 10.1038/s41598-020-58050-2. Sci Rep. 2020. PMID: 32024875 Free PMC article.
-
Isolation of T cells from mouse oral tissues.Biol Proced Online. 2014 Mar 10;16(1):4. doi: 10.1186/1480-9222-16-4. Biol Proced Online. 2014. PMID: 24612879 Free PMC article.
References
-
- Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006;126:663–676. - PubMed
-
- Liu H, Zhu F, Yong J, Zhang P, Hou P, et al. Generation of induced pluripotent stem cells from adult rhesus monkey fibroblasts. Cell Stem Cell. 2008;3:587–590. - PubMed
-
- Shimada H, Nakada A, Hashimoto Y, Shigeno K, Shionoya Y, et al. Generation of canine induced pluripotent stem cells by retroviral transduction and chemical inhibitors. Mol Reprod Dev. 2010;77:2. - PubMed
-
- Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131:861–872. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
