

Involvement of the melanocortin-1 receptor in acute pain and pain of inflammatory but not neuropathic origin
- PMID: 20856883
- PMCID: PMC2938350
- DOI: 10.1371/journal.pone.0012498
Involvement of the melanocortin-1 receptor in acute pain and pain of inflammatory but not neuropathic origin
Abstract
Background: Response to painful stimuli is susceptible to genetic variation. Numerous loci have been identified which contribute to this variation, one of which, MC1R, is better known as a gene involved in mammalian hair colour. MC1R is a G protein-coupled receptor expressed in melanocytes and elsewhere and mice lacking MC1R have yellow hair, whilst humans with variant MC1R protein have red hair. Previous work has found differences in acute pain perception, and response to analgesia in mice and humans with mutations or variants in MC1R.
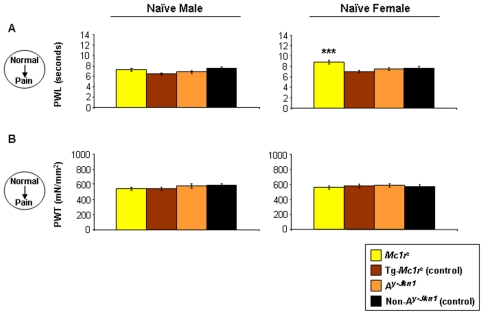
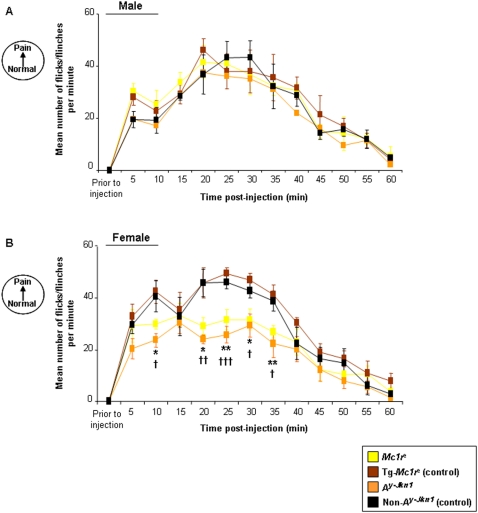
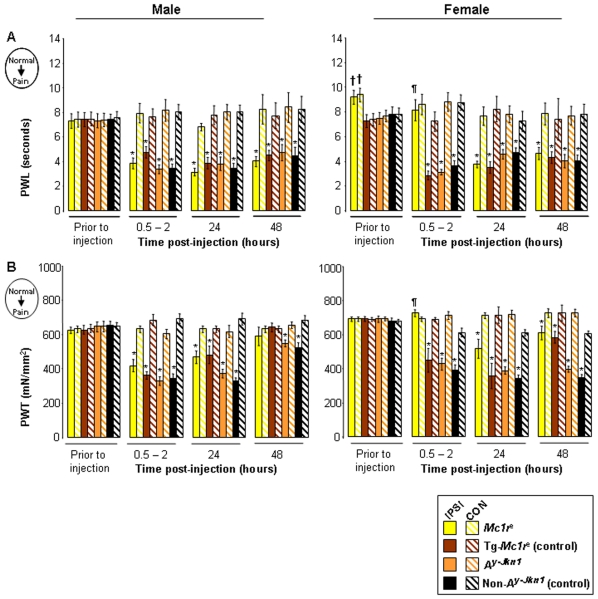
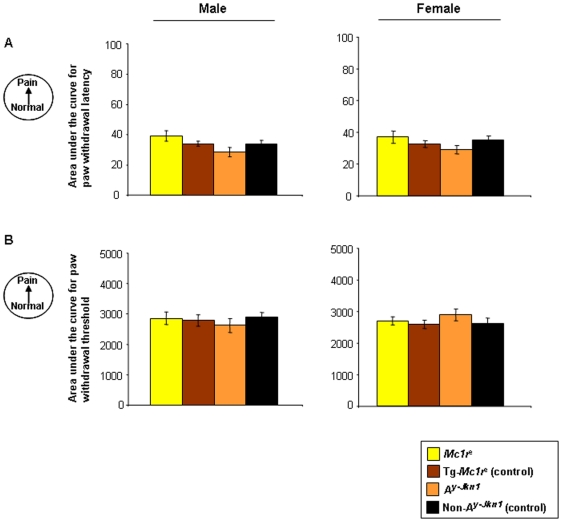
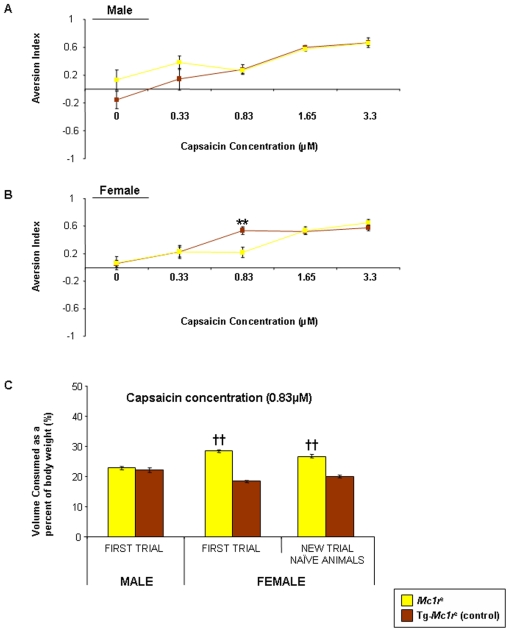
Methodology and principal findings: We have tested responses to noxious and non-noxious stimuli in mutant mice which lack MC1R, or which overexpress an endogenous antagonist of the receptor, as well as controls. We have also examined the response of these mice to inflammatory pain, assessing the hyperalgesia and allodynia associated with persistent inflammation, and their response to neuropathic pain. Finally we tested by a paired preference paradigm their aversion to oral administration of capsaicin, which activates the noxious heat receptor TRPV1. Female mice lacking MC1R showed increased tolerance to noxious heat and no alteration in their response to non-noxious mechanical stimuli. MC1R mutant females, and females overexpressing the endogenous MC1R antagonist, agouti signalling protein, had a reduced formalin-induced inflammatory pain response, and a delayed development of inflammation-induced hyperalgesia and allodynia. In addition they had a decreased aversion to capsaicin at moderate concentrations. Male mutant mice showed no difference from their respective controls. Mice of either sex did not show any effect of mutant genotype on neuropathic pain.
Conclusions: We demonstrate a sex-specific role for MC1R in acute noxious thermal responses and pain of inflammatory origin.
Conflict of interest statement
Figures
Similar articles
-
Relationship between the melanocortin-1 receptor (MC1R) variant R306ter and physiological responses to mechanical or thermal stimuli in Labrador Retriever dogs.Vet Anaesth Analg. 2017 Mar;44(2):370-374. doi: 10.1016/j.vaa.2016.05.003. Epub 2017 Jan 11. Vet Anaesth Analg. 2017. PMID: 28214221 Free PMC article.
-
Melanocortin-1 receptor gene variants affect pain and mu-opioid analgesia in mice and humans.J Med Genet. 2005 Jul;42(7):583-7. doi: 10.1136/jmg.2004.027698. J Med Genet. 2005. PMID: 15994880 Free PMC article.
-
Detangling red hair from pain: phenotype-specific contributions from different genetic variants in melanocortin-1 receptor.Pain. 2020 May;161(5):938-948. doi: 10.1097/j.pain.0000000000001780. Pain. 2020. PMID: 31834199 Free PMC article.
-
Melanocortins and the melanocortin 1 receptor, moving translationally towards melanoma prevention.Arch Biochem Biophys. 2014 Dec 1;563:4-12. doi: 10.1016/j.abb.2014.07.002. Epub 2014 Jul 11. Arch Biochem Biophys. 2014. PMID: 25017567 Review.
-
MC1R and the response of melanocytes to ultraviolet radiation.Mutat Res. 2005 Apr 1;571(1-2):133-52. doi: 10.1016/j.mrfmmm.2004.09.014. Epub 2005 Jan 26. Mutat Res. 2005. PMID: 15748644 Review.
Cited by
-
Bench-top to clinical therapies: A review of melanocortin ligands from 1954 to 2016.Biochim Biophys Acta Mol Basis Dis. 2017 Oct;1863(10 Pt A):2414-2435. doi: 10.1016/j.bbadis.2017.03.020. Epub 2017 Mar 29. Biochim Biophys Acta Mol Basis Dis. 2017. PMID: 28363699 Free PMC article. Review.
-
Pharmacogenomics of Pain Management: The Impact of Specific Biological Polymorphisms on Drugs and Metabolism.Curr Oncol Rep. 2020 Feb 6;22(2):18. doi: 10.1007/s11912-020-0865-4. Curr Oncol Rep. 2020. PMID: 32030524 Review.
-
Relationship between the melanocortin-1 receptor (MC1R) variant R306ter and physiological responses to mechanical or thermal stimuli in Labrador Retriever dogs.Vet Anaesth Analg. 2017 Mar;44(2):370-374. doi: 10.1016/j.vaa.2016.05.003. Epub 2017 Jan 11. Vet Anaesth Analg. 2017. PMID: 28214221 Free PMC article.
-
Preliminary investigation of potential links between pigmentation variants and opioid analgesic effectiveness in horses during cerebrospinal fluid centesis.BMC Vet Res. 2024 Jul 12;20(1):311. doi: 10.1186/s12917-024-04139-z. BMC Vet Res. 2024. PMID: 38997753 Free PMC article.
-
Machine-learned analysis of the association of next-generation sequencing-based human TRPV1 and TRPA1 genotypes with the sensitivity to heat stimuli and topically applied capsaicin.Pain. 2018 Jul;159(7):1366-1381. doi: 10.1097/j.pain.0000000000001222. Pain. 2018. PMID: 29596157 Free PMC article.
References
-
- Julius D, Basbaum AI. Molecular mechanisms of nociception. Nature. 2001;413:203–210. - PubMed
-
- Dyck PJ. Neuronal atrophy and degeneration predominantly affecting peripheral sensory and autonomic neurons. In: Dyck PJ, Thomas PK, Griffi JW, Low PW, Poduslo JF, editors. Peripheral Neuropathy. Philadelphia: Saunders; 1993. pp. 1065–1093.
-
- Craft RM. Sex differences in opioid analgesia: “from mouse to man”. Clin J Pain. 2003;19:175–186. - PubMed
-
- Wiesenfeld-Hallin Z. Sex differences in pain perception. Gend Med. 2005;2:137–145. - PubMed
-
- Berkley KJ. Sex differences in pain. Behav Brain Sci. 1997;20:371–380. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
