

The cercal organ may provide singing tettigoniids a backup sensory system for the detection of eavesdropping bats
- PMID: 20856887
- PMCID: PMC2938355
- DOI: 10.1371/journal.pone.0012698
The cercal organ may provide singing tettigoniids a backup sensory system for the detection of eavesdropping bats
Abstract
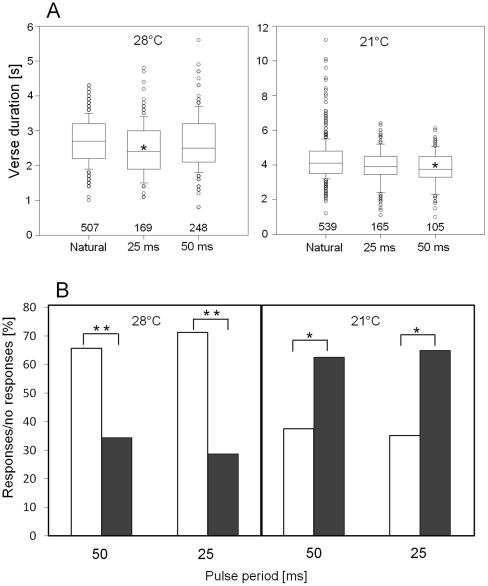
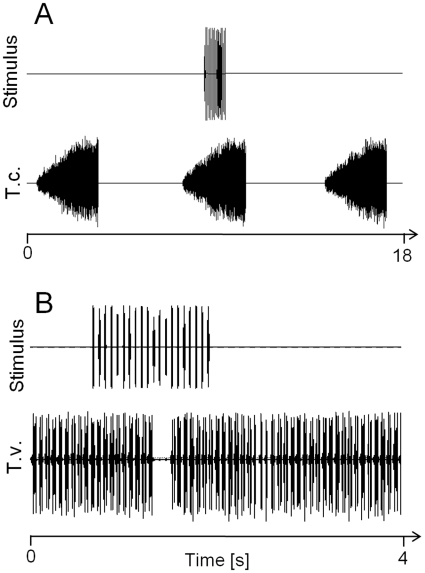
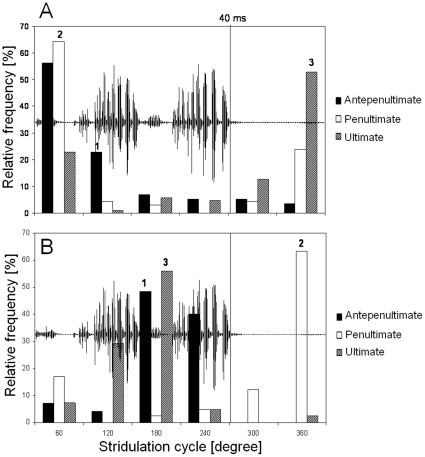
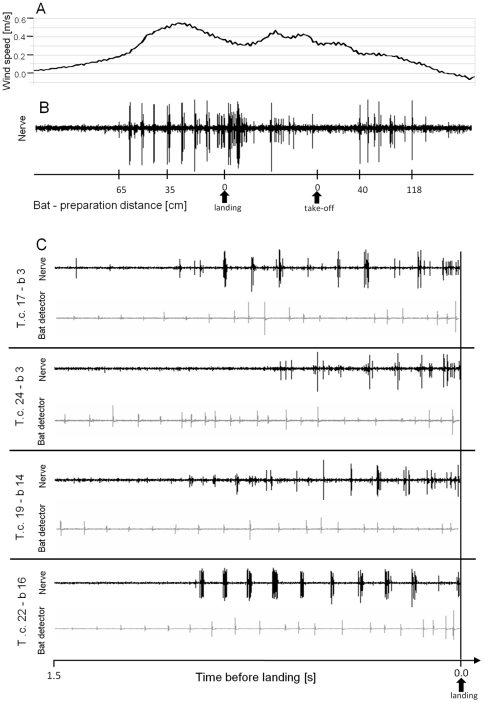
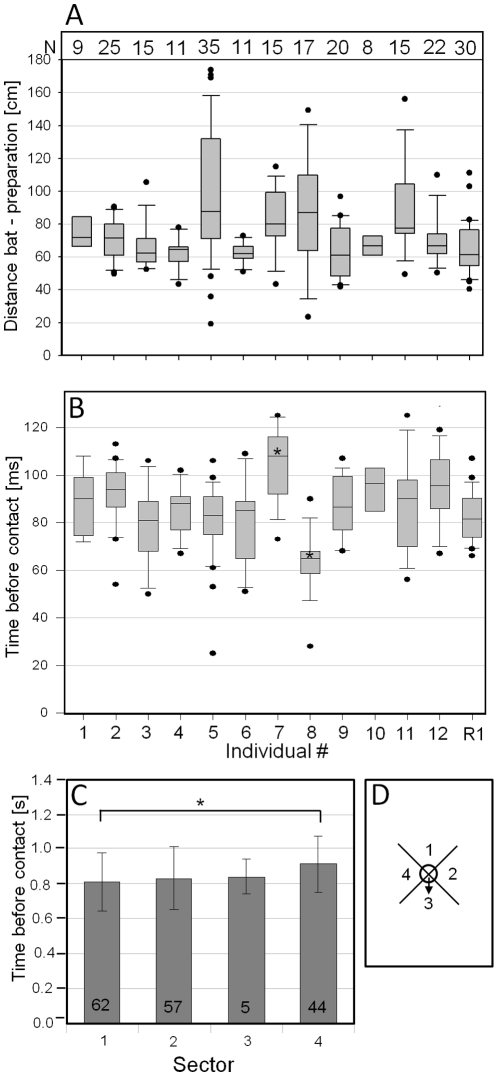
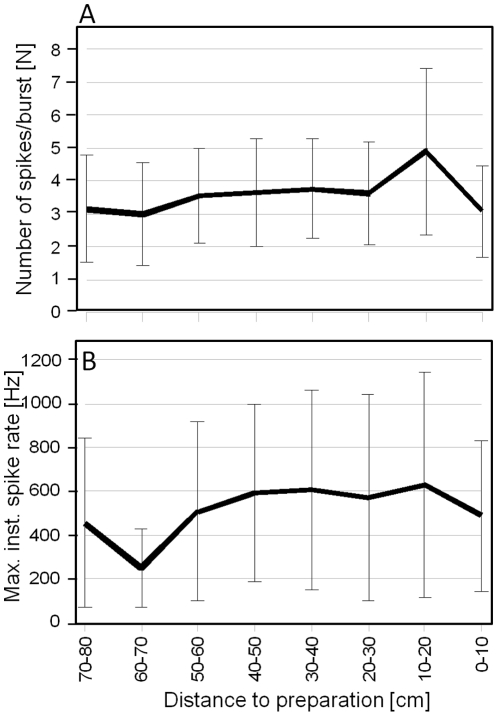
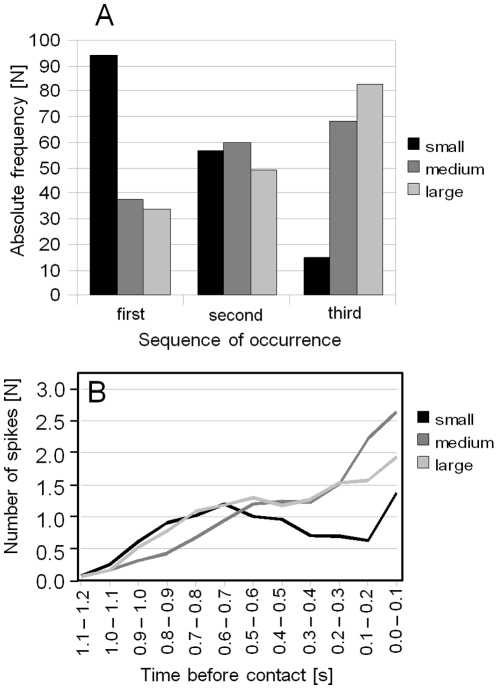
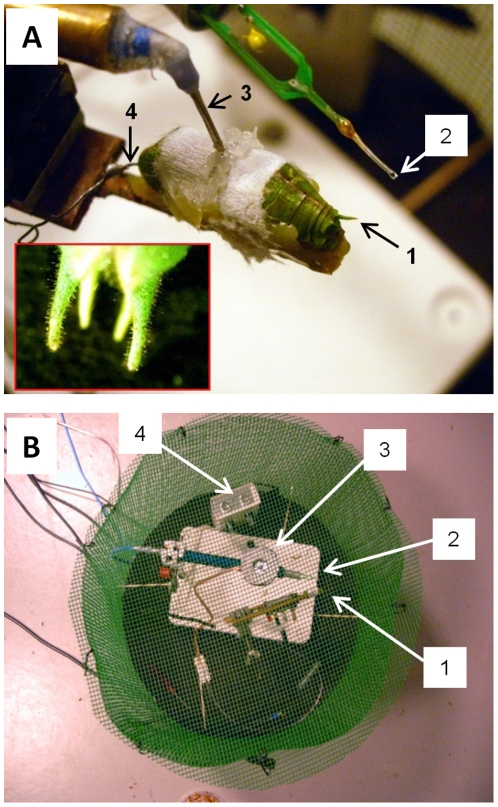
Conspicuous signals, such as the calling songs of tettigoniids, are intended to attract mates but may also unintentionally attract predators. Among them bats that listen to prey-generated sounds constitute a predation pressure for many acoustically communicating insects as well as frogs. As an adaptation to protect against bat predation many insect species evolved auditory sensitivity to bat-emitted echolocation signals. Recently, the European mouse-eared bat species Myotis myotis and M. blythii oxygnathus were found to eavesdrop on calling songs of the tettigoniid Tettigonia cantans. These gleaning bats emit rather faint echolocation signals when approaching prey and singing insects may have difficulty detecting acoustic predator-related signals. The aim of this study was to determine (1) if loud self-generated sound produced by European tettigoniids impairs the detection of pulsed ultrasound and (2) if wind-sensors on the cercal organ function as a sensory backup system for bat detection in tettigoniids. We addressed these questions by combining a behavioral approach to study the response of two European tettigoniid species to pulsed ultrasound, together with an electrophysiological approach to record the activity of wind-sensitive interneurons during real attacks of the European mouse-eared bat species Myotis myotis. Results showed that singing T. cantans males did not respond to sequences of ultrasound pulses, whereas singing T. viridissima did respond with predominantly brief song pauses when ultrasound pulses fell into silent intervals or were coincident with the production of soft hemi-syllables. This result, however, strongly depended on ambient temperature with a lower probability for song interruption observable at 21°C compared to 28°C. Using extracellular recordings, dorsal giant interneurons of tettigoniids were shown to fire regular bursts in response to attacking bats. Between the first response of wind-sensitive interneurons and contact, a mean time lag of 860 ms was found. This time interval corresponds to a bat-to-prey distance of ca. 72 cm. This result demonstrates the efficiency of the cercal system of tettigoniids in detecting attacking bats and suggests this sensory system to be particularly valuable for singing insects that are targeted by eavesdropping bats.
Conflict of interest statement
Figures

References
-
- Hoy RR. The evolution of hearing in insects as an adaptation to predation from bats. In: Webster DB, Fay RR, Popper AN, editors. The evolutionary biology of hearing. Berlin Heidelberg New York: Springer; 1992. pp. 115–129.
-
- Fullard JH, Yack JE. The evolutionary biology of insect hearing. Trends Ecol Evol. 1993;8:248–252. - PubMed
-
- Hoy RR, Robert D. Tympanal hearing in insects. Annu Rev Entomol. 1996;41:433–450. - PubMed
-
- Yager D. Structure, development, and evolution of insect auditory systems. Micros Res Techn. 1999;47:380–400. - PubMed
-
- Triblehorn JD, Yager DD. Wind generated by an attacking bat: anemometric measurements and detection by the praying mantis cercal system. J Exp Biol. 2006;209:1430–1440. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
