

Cone and rod photoreceptor transplantation in models of the childhood retinopathy Leber congenital amaurosis using flow-sorted Crx-positive donor cells
- PMID: 20858907
- PMCID: PMC2972691
- DOI: 10.1093/hmg/ddq378
Cone and rod photoreceptor transplantation in models of the childhood retinopathy Leber congenital amaurosis using flow-sorted Crx-positive donor cells
Abstract
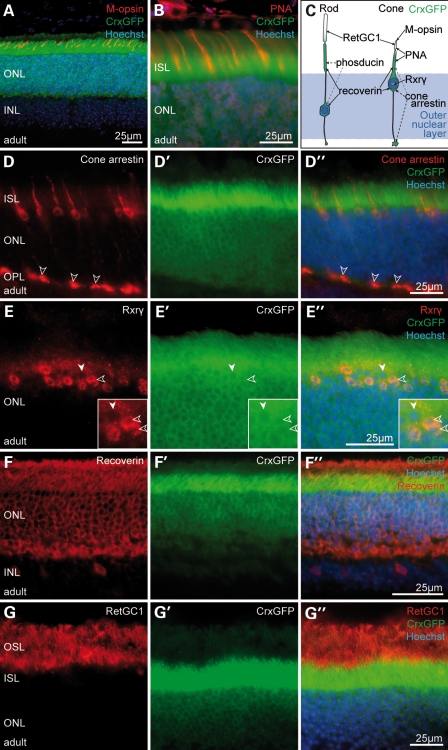
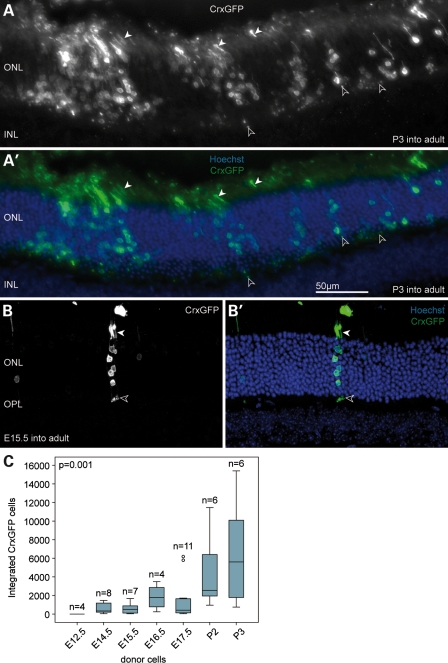
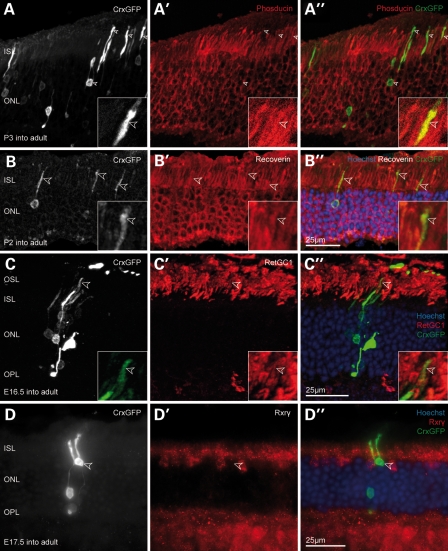
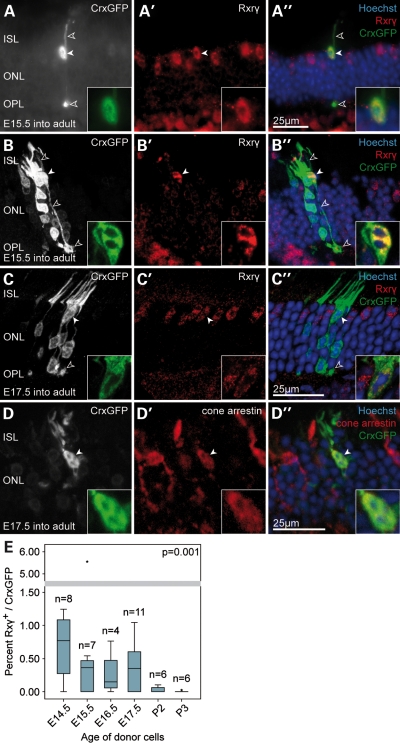
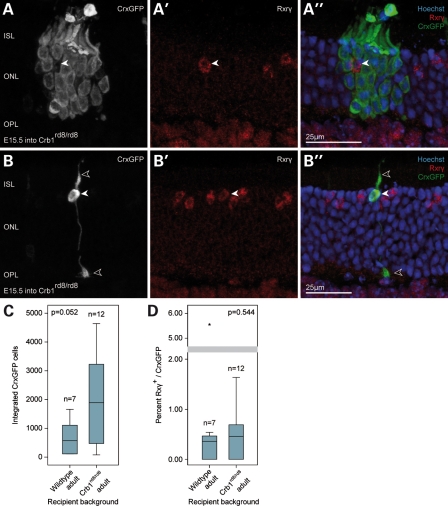
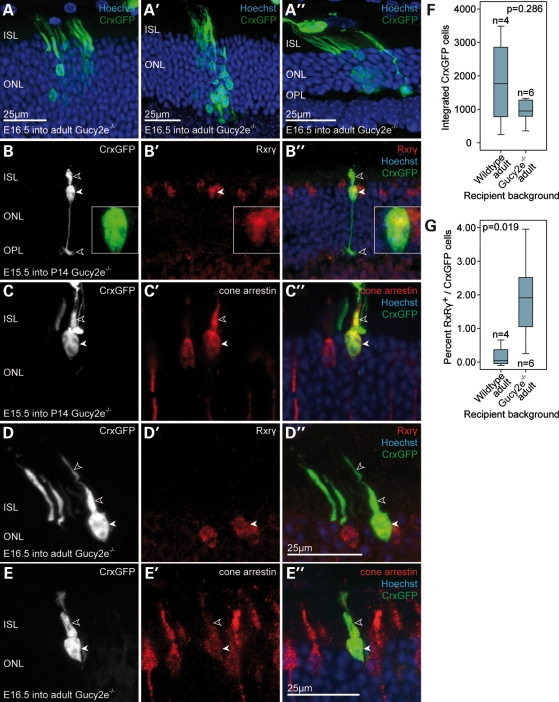
Retinal degenerative disease causing loss of photoreceptor cells is the leading cause of untreatable blindness in the developed world, with inherited degeneration affecting 1 in 3000 people. Visual acuity deteriorates rapidly once the cone photoreceptors die, as these cells provide daylight and colour vision. Here, in proof-of-principle experiments, we demonstrate the feasibility of cone photoreceptor transplantation into the wild-type and degenerating retina of two genetic models of Leber congenital amaurosis, the Crb1(rd8/rd8) and Gucy2e(-/-) mouse. Crx-expressing cells were flow-sorted from the developing retina of CrxGFP transgenic mice and transplanted into adult recipient retinae; CrxGFP is a marker of cone and rod photoreceptor commitment. Only the embryonic-stage Crx-positive donor cells integrated within the outer nuclear layer of the recipient and differentiated into new cones, whereas postnatal cells generated a 10-fold higher number of rods compared with embryonic-stage donors. New cone photoreceptors displayed unambiguous morphological cone features and expressed mature cone markers. Importantly, we found that the adult environment influences the number of integrating cones and favours rod integration. New cones and rods were observed in ratios similar to that of the host retina (1:35) even when the transplanted population consisted primarily of cone precursors. Cone integration efficiency was highest in the cone-deficient Gucy2e(-/-) retina suggesting that cone depletion creates a more optimal environment for cone transplantation. This is the first comprehensive study demonstrating the feasibility of cone transplantation into the adult retina. We conclude that flow-sorted embryonic-stage Crx-positive donor cells have the potential to replace lost cones, as well as rods, an important requirement for retinal disease therapy.
Figures
References
-
- Morrow E.M., Furukawa T., Cepko C.L. Vertebrate photoreceptor cell development and disease. Trends Cell Biol. 1998;8:353–358. doi:10.1016/S0962-8924(98)01341-5. - DOI - PubMed
-
- Hartong D.T., Berson E.L., Dryja T.P. Retinitis pigmentosa. Lancet. 2006;368:1795–1809. doi:10.1016/S0140-6736(06)69740-7. - DOI - PubMed
-
- Lamba D.A., Karl M.O., Reh T.A. Strategies for retinal repair: cell replacement and regeneration. Prog. Brain Res. 2009;175:23–31. doi:10.1016/S0079-6123(09)17502-7. - DOI - PubMed
-
- West E.L., Pearson R.A., MacLaren R.E., Sowden J.C., Ali R.R. Cell transplantation strategies for retinal repair. Prog. Brain Res. 2009;175:3–21. doi:10.1016/S0079-6123(09)17501-5. - DOI - PMC - PubMed
-
- MacLaren R.E., Pearson R.A., MacNeil A., Douglas R.H., Salt T.E., Akimoto M., Swaroop A., Sowden J.C., Ali R.R. Retinal repair by transplantation of photoreceptor precursors. Nature. 2006;444:203–207. doi:10.1038/nature05161. - DOI - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
