

Unusual development of light-reflecting pigment cells in intact and regenerating tail in the periodic albino mutant of Xenopus laevis
- PMID: 20859642
- PMCID: PMC2948654
- DOI: 10.1007/s00441-010-1042-0
Unusual development of light-reflecting pigment cells in intact and regenerating tail in the periodic albino mutant of Xenopus laevis
Abstract
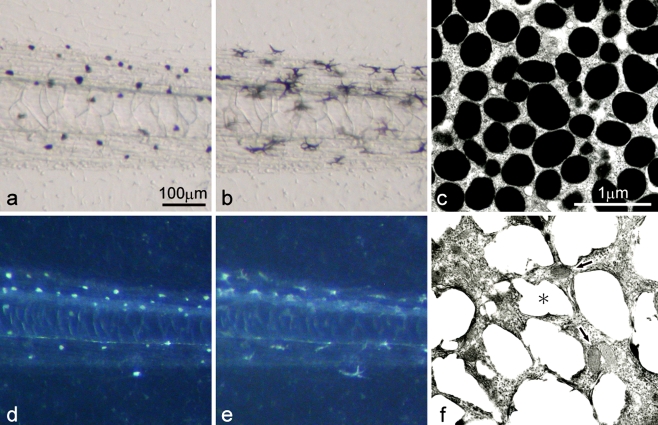
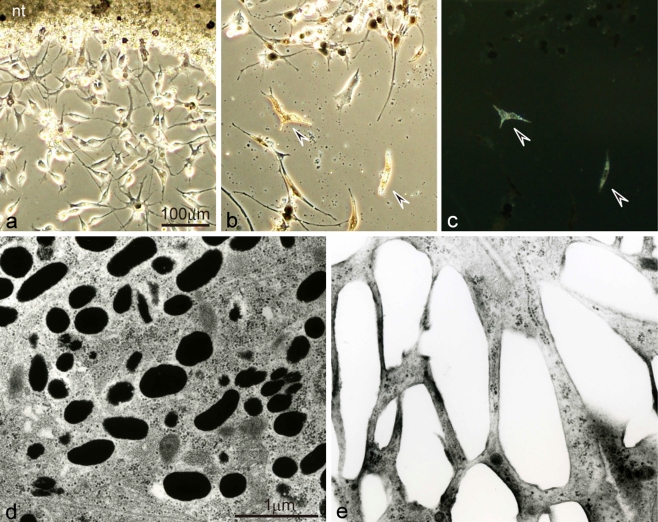
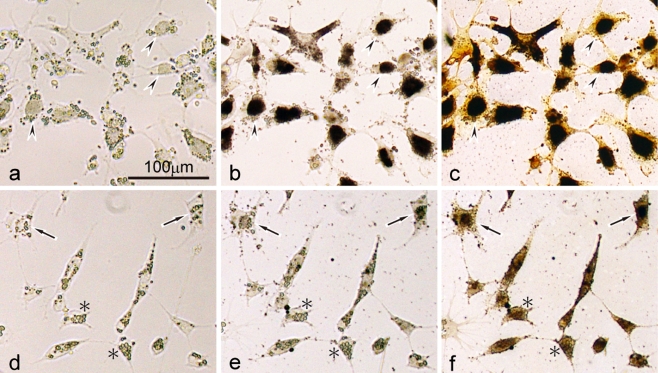
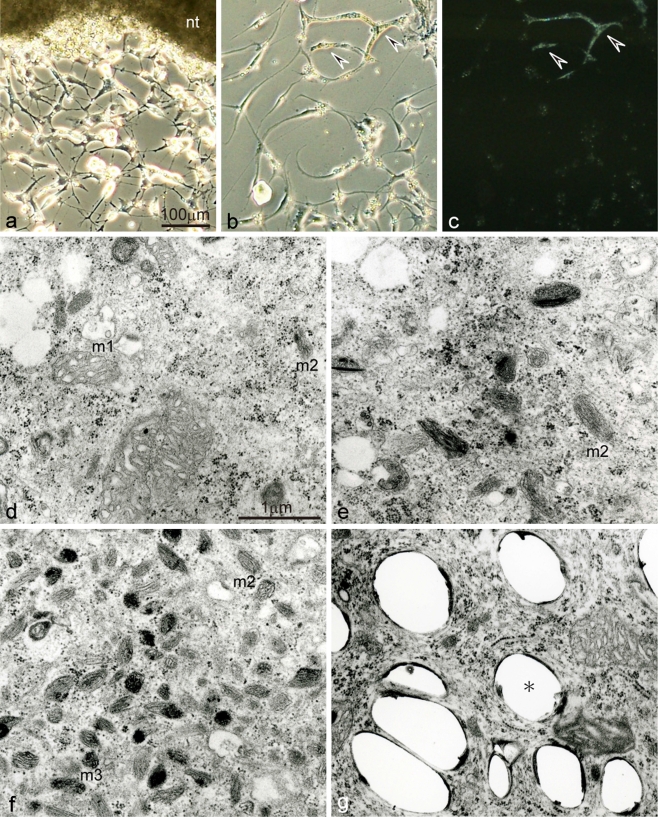
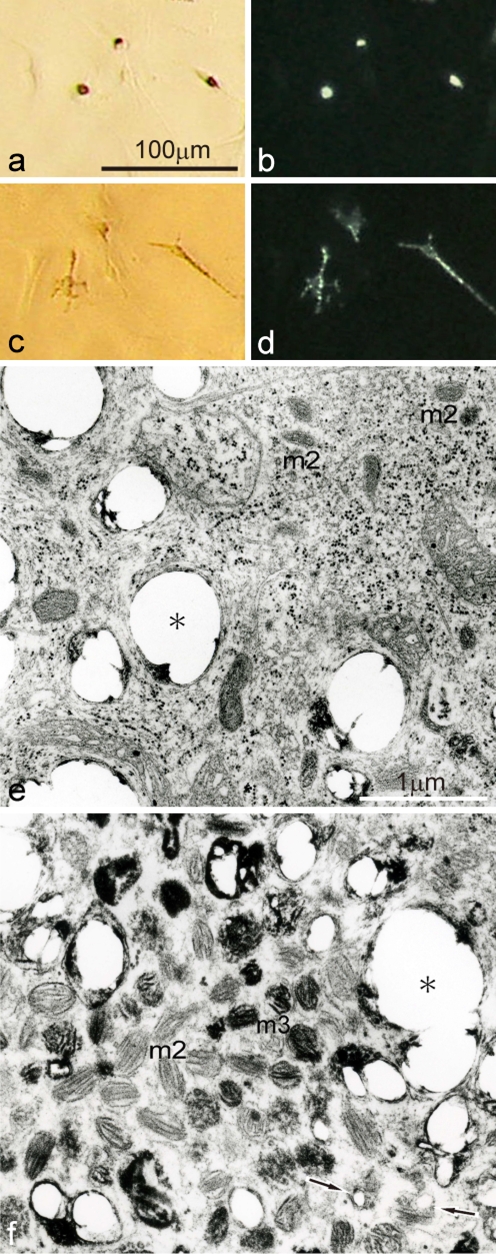
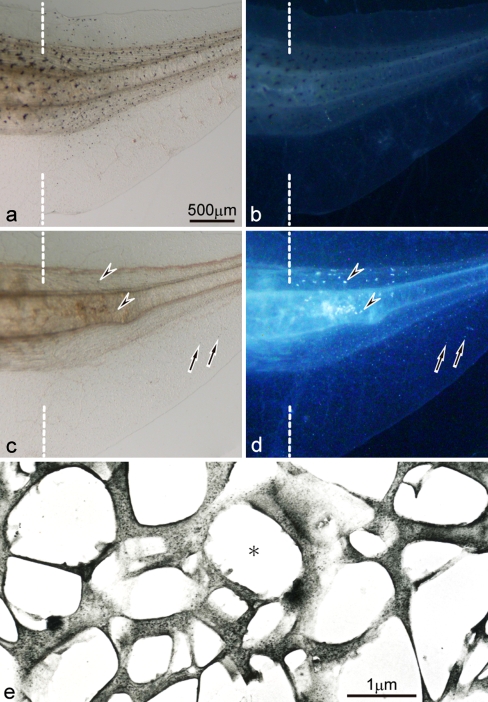
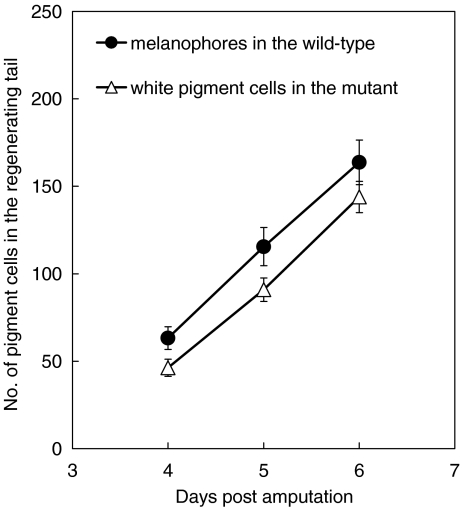
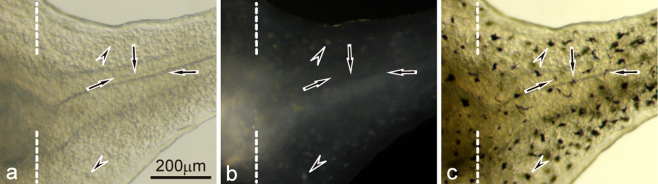
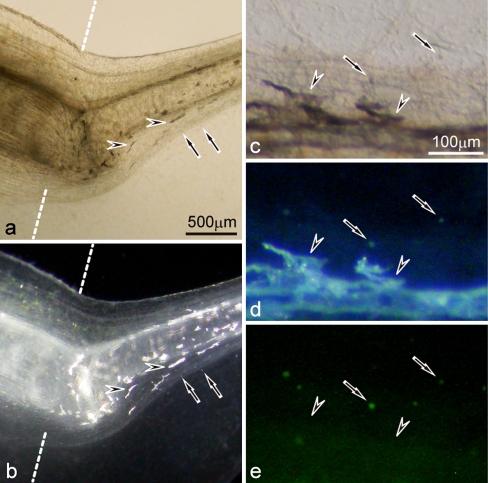
Unusual light-reflecting pigment cells, "white pigment cells", specifically appear in the periodic albino mutant (a(p) /a(p)) of Xenopus laevis and localize in the same place where melanophores normally differentiate in the wild-type. The mechanism responsible for the development of unusual pigment cells is unclear. In this study, white pigment cells in the periodic albino were compared with melanophores in the wild-type, using a cell culture system and a tail-regenerating system. Observations of both intact and cultured cells demonstrate that white pigment cells are unique in (1) showing characteristics of melanophore precursors at various stages of development, (2) accumulating reflecting platelets characteristic of iridophores, and (3) exhibiting pigment dispersion in response to α-melanocyte stimulating hormone (α-MSH) in the same way that melanophores do. When a tadpole tail is amputated, a functionally competent new tail is regenerated. White pigment cells appear in the mutant regenerating tail, whereas melanophores differentiate in the wild-type regenerating tail. White pigment cells in the mutant regenerating tail are essentially similar to melanophores in the wild-type regenerating tail with respect to their localization, number, and response to α-MSH. In addition to white pigment cells, iridophores which are never present in the intact tadpole tail appear specifically in the somites near the amputation level in the mutant regenerating tail. Iridophores are distinct from white pigment cells in size, shape, blue light-induced fluorescence, and response to α-MSH. These findings strongly suggest that white pigment cells in the mutant arise from melanophore precursors and accumulate reflecting platelets characteristic of iridophores.
Figures
References
-
- Bagnara JT. Hypophysectomy and the tail darkening reaction in Xenopus. Proc Soc Exp Biol Med. 1957;94:572–575. - PubMed
-
- Bagnara JT. Tail melanophores of Xenopus in normal development and regeneration. Biol Bull. 1960;118:1–8. doi: 10.2307/1539050. - DOI
-
- Bagnara JT. The neural crest as a source of stem cells. In: Maderson P, editor. Developmental and evolutionary aspects of the neural crest. New York: Wiley; 1987. pp. 57–87.
-
- Bagnara JT. Comparative anatomy and physiology of pigment cells in nonmammalian tissues. In: Nordlund JJ, Boissy RE, Hearing VJ, King RA, Ortonne J-P, editors. The pigmentary system: physiology and pathophysiology. New York: Oxford University Press; 1998. pp. 9–40.
-
- Bagnara JT, Hadley ME. Chromatophores and color change: the comparative physiology of animal pigmentation. Englewood Cliffs, NJ: Prentice-Hall; 1973.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
