

Aquaporins: relevance to cerebrospinal fluid physiology and therapeutic potential in hydrocephalus
- PMID: 20860832
- PMCID: PMC2949735
- DOI: 10.1186/1743-8454-7-15
Aquaporins: relevance to cerebrospinal fluid physiology and therapeutic potential in hydrocephalus
Abstract
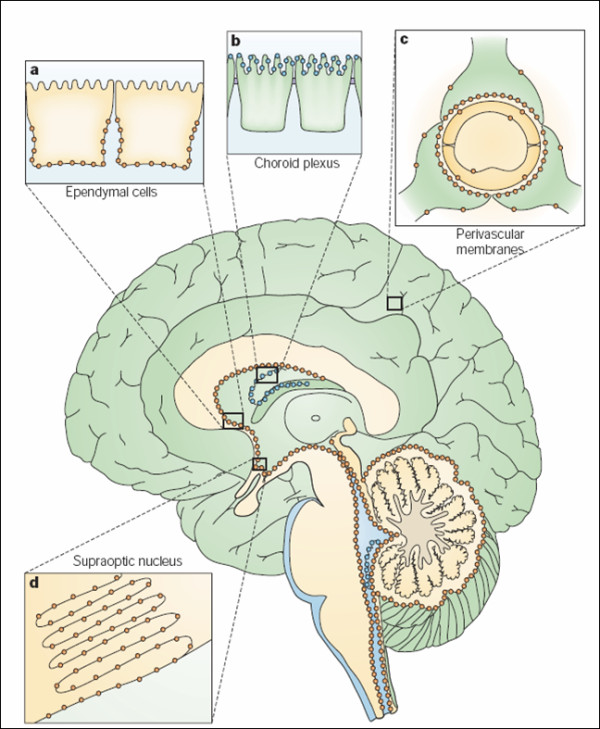
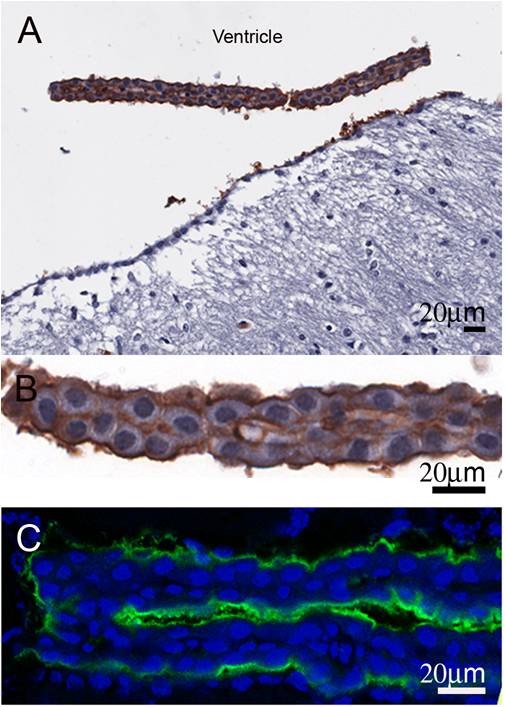
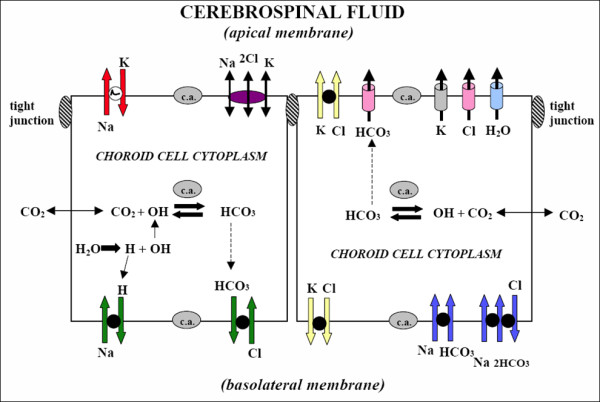
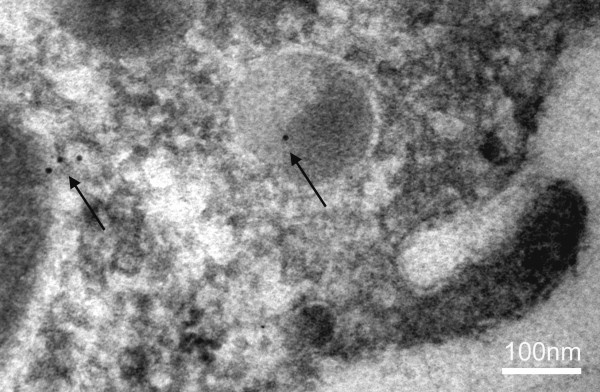
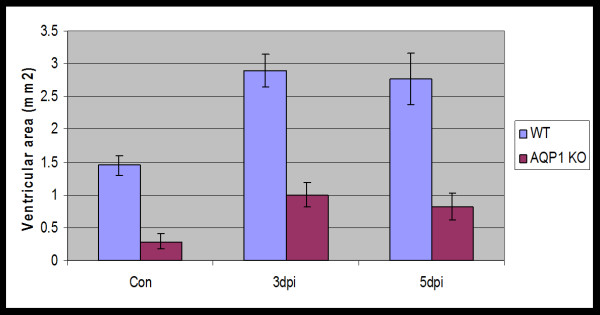
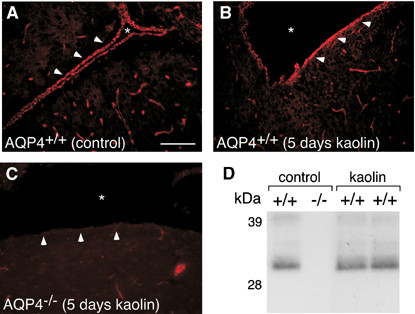
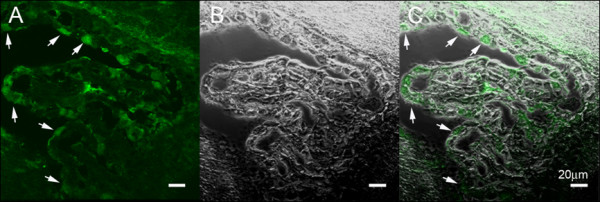
The discovery of a family of membrane water channel proteins called aquaporins, and the finding that aquaporin 1 was located in the choroid plexus, has prompted interest in the role of aquaporins in cerebrospinal fluid (CSF) production and consequently hydrocephalus. While the role of aquaporin 1 in choroidal CSF production has been demonstrated, the relevance of aquaporin 1 to the pathophysiology of hydrocephalus remains debated. This has been further hampered by the lack of a non-toxic specific pharmacological blocking agent for aquaporin 1. In recent times aquaporin 4, the most abundant aquaporin within the brain itself, which has also been shown to have a role in brain water physiology and relevance to brain oedema in trauma and tumours, has become an alternative focus of attention for hydrocephalus research. This review summarises current knowledge and concepts in relation to aquaporins, specifically aquaporin 1 and 4, and hydrocephalus. It also examines the relevance of aquaporins as potential therapeutic targets in hydrocephalus and other CSF circulation disorders.
Figures

References
LinkOut - more resources
Full Text Sources
