

A unique redox-sensing sensor II motif in TorsinA plays a critical role in nucleotide and partner binding
- PMID: 20861018
- PMCID: PMC2988333
- DOI: 10.1074/jbc.M110.123471
A unique redox-sensing sensor II motif in TorsinA plays a critical role in nucleotide and partner binding
Abstract
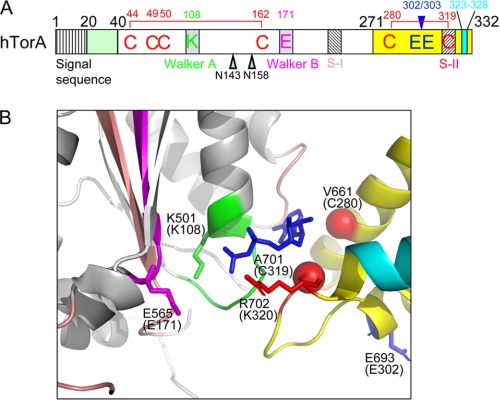
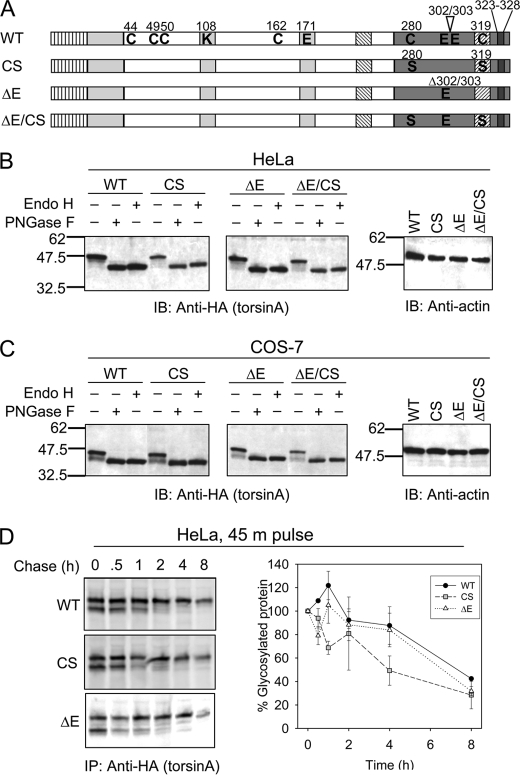
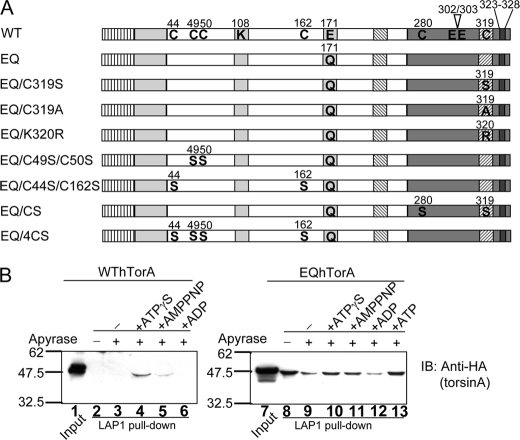
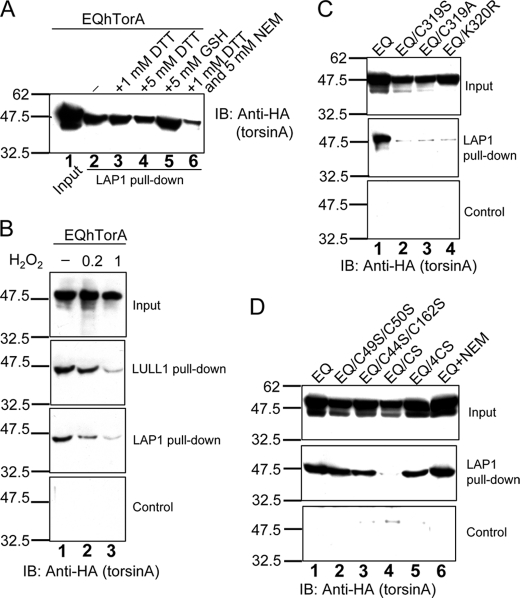
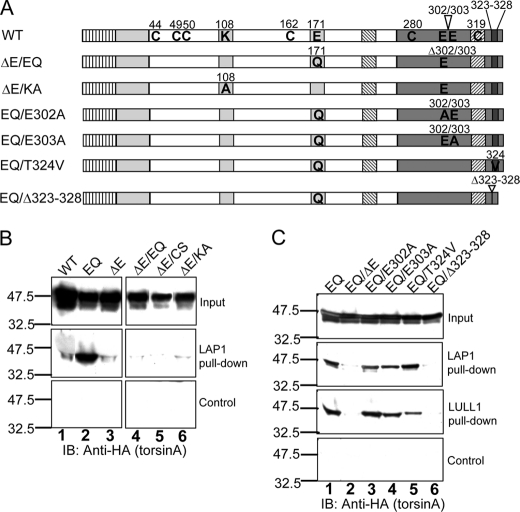
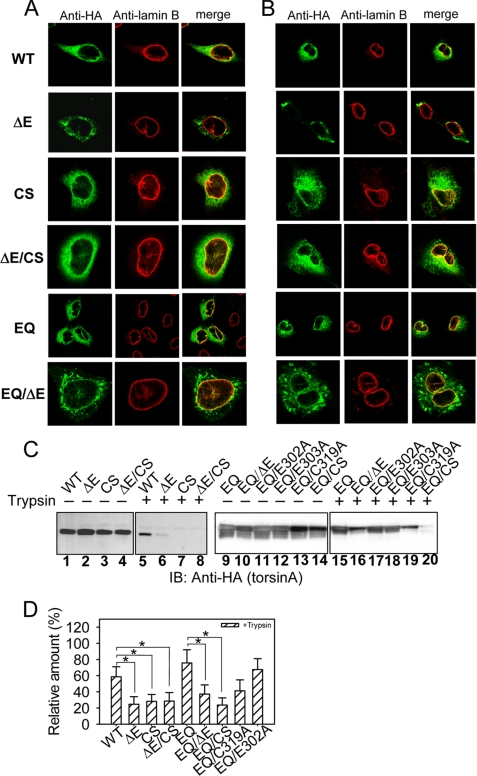
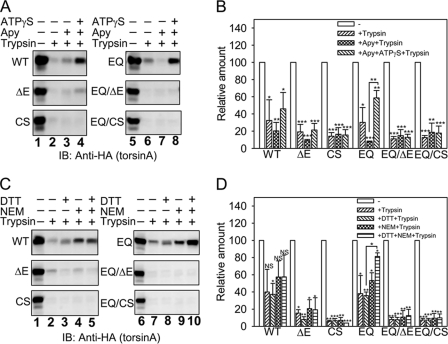
Early onset dystonia is commonly associated with the deletion of one of a pair of glutamate residues (ΔE302/303) near the C terminus of torsinA, a member of the AAA+ protein family (ATPases associated with a variety of cellular activities) located in the endoplasmic reticulum lumen. The functional consequences of the disease-causing mutation, ΔE, are not currently understood. By contrast to other AAA+ proteins, torsin proteins contain two conserved cysteine residues in the C-terminal domain, one of which is located in the nucleotide sensor II motif. Depending on redox status, an ATP hydrolysis mutant of torsinA interacts with lamina-associated polypeptide 1 (LAP1) and lumenal domain like LAP1 (LULL1). Substitution of the cysteine in sensor II diminishes the redox-regulated interaction of torsinA with these substrates. Significantly, the dystonia-causing mutation, ΔE, alters the ability of torsinA to mediate the redox-regulated interactions with LAP1 and LULL1. Limited proteolysis experiments reveal redox- and mutation-dependent changes in the local conformation of torsinA as a function of nucleotide binding. These results indicate that the cysteine-containing sensor II plays a critical role in redox sensing and the nucleotide and partner binding functions of torsinA and suggest that loss of this function of torsinA contributes to the development of DYT1 dystonia.
Figures
References
-
- Ozelius L. J., Hewett J. W., Page C. E., Bressman S. B., Kramer P. L., Shalish C., de Leon D., Brin M. F., Raymond D., Corey D. P., Fahn S., Risch N. J., Buckler A. J., Gusella J. F., Breakefield X. O. (1997) Nat. Genet. 17, 40–48 - PubMed
-
- Breakefield X. O., Blood A. J., Li Y., Hallett M., Hanson P. I., Standaert D. G. (2008) Nat. Rev. Neurosci. 9, 222–234 - PubMed
-
- Neuwald A. F., Aravind L., Spouge J. L., Koonin E. V. (1999) Genome Res. 9, 27–43 - PubMed
-
- Hanson P. I., Whiteheart S. W. (2005) Nat. Rev. Mol. Cell Biol. 6, 519–529 - PubMed
-
- Doyle S. M., Wickner S. (2009) Trends Biochem. Sci 34, 40–48 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
