

Within and transgenerational immune priming in an insect to a DNA virus
- PMID: 20861049
- PMCID: PMC3049047
- DOI: 10.1098/rspb.2010.1517
Within and transgenerational immune priming in an insect to a DNA virus
Abstract
Invertebrates mount a sophisticated immune response with the potential to exhibit a form of immune memory through 'priming'. Increased immune protection following early exposure to bacteria has been found both later in life (within generation priming) and in the next generation (transgeneration priming) in a number of invertebrates. However, it is unclear how general immune priming is and whether immune priming occurs in response to different parasites, including viruses. Here, using Plodia interpuctella (Lepidoptera) and its natural DNA virus, Plodia interpunctella granulosis virus, we find evidence for both within generation and transgeneration immune priming. Individuals previously exposed to low doses of virus, as well as the offspring of exposed individuals, are subsequently less susceptible to viral challenge. Relatively little is known about the mechanisms that underpin viral immunity but it is probable that the viral immune response is somewhat different to that of bacteria. We show that immune priming may, however, be a characteristic of both responses, mediated through different mechanisms, suggesting that immune memory may be a general phenomenon of insect immunity. This is important because immune priming may influence both host-parasite population and evolutionary dynamics.
Figures
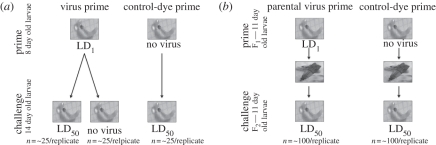
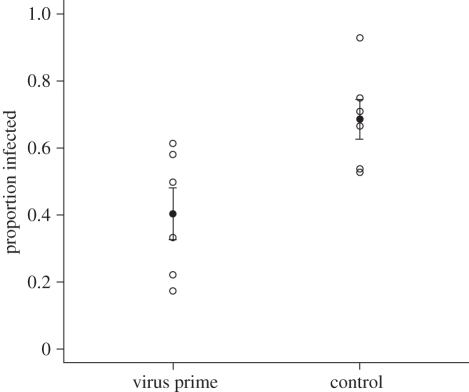
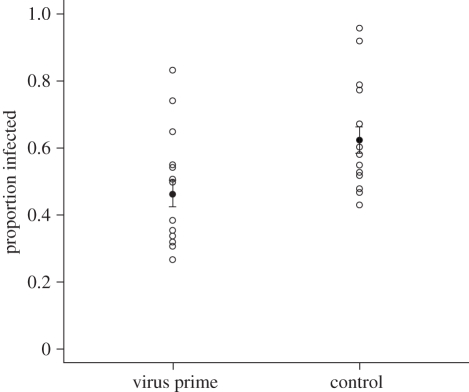
References
-
- Anderson R. M., May R. M. 1981. The population-dynamics of micro-parasites and their invertebrate hosts. Phil. Trans. R. Soc. Lond. B 291, 451–524 10.1098/rstb.1981.0005 (doi:10.1098/rstb.1981.0005) - DOI
-
- Boots M., Begon M. 1993. Trade-offs with resistance to a granulosis-virus in the Indian meal moth, examined by a laboratory evolution experiment. Funct. Ecol. 7, 528–534 10.2307/2390128 (doi:10.2307/2390128) - DOI
-
- Hudson P. J., Dobson A. P., Newborn D. 1998. Prevention of population cycles by parasite removal. Science 282, 2256–2258 10.1126/science.282.5397.2256 (doi:10.1126/science.282.5397.2256) - DOI - PubMed
-
- Pedersen A. B., Greives T. J. 2008. The interaction of parasites and resources cause crashes in a wild mouse population. J. Anim. Ecol. 77, 370–377 10.1111/j.1365-2656.2007.01321.x (doi:10.1111/j.1365-2656.2007.01321.x) - DOI - PubMed
-
- Hatcher M. J., Dick J. T. A., Dunn A. M. 2006. How parasites affect interactions between competitors and predators. Ecol. Lett. 9, 1253–1271 10.1111/j.1461-0248.2006.00964.x (doi:10.1111/j.1461-0248.2006.00964.x) - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources