

A double mutation in glycoprotein gB compensates for ineffective gD-dependent initiation of herpes simplex virus type 1 infection
- PMID: 20861246
- PMCID: PMC2976401
- DOI: 10.1128/JVI.01633-10
A double mutation in glycoprotein gB compensates for ineffective gD-dependent initiation of herpes simplex virus type 1 infection
Abstract
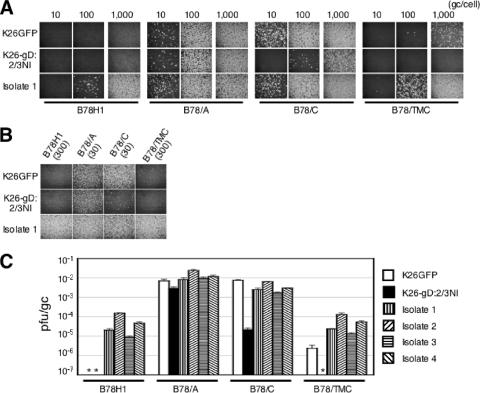
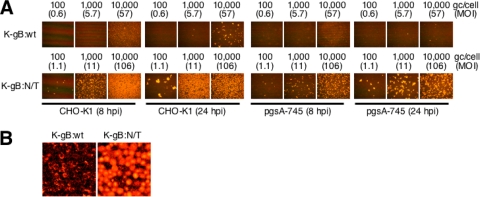
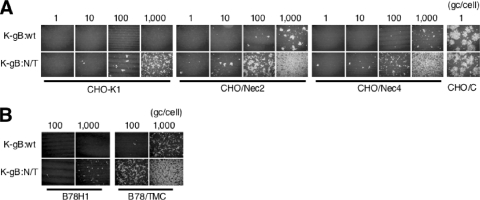
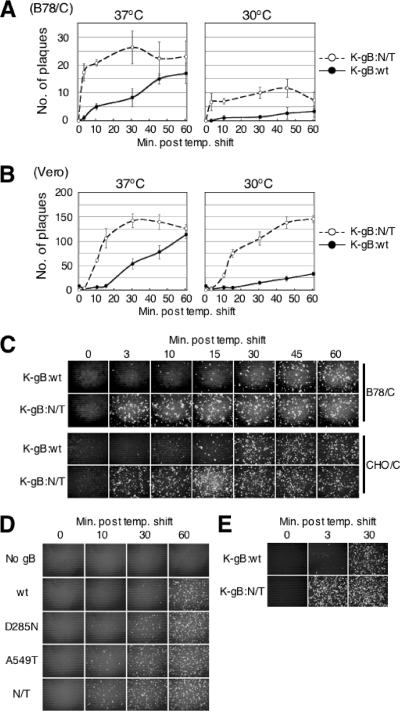
Herpes simplex virus (HSV) entry into cells is triggered by the binding of envelope glycoprotein D (gD) to a specific receptor, such as nectin-1 or herpesvirus entry mediator (HVEM), resulting in activation of the fusion effectors gB and gH and virus penetration. Here we report the identification of a hyperactive gB allele, D285N/A549T, selected by repeat passage of a gD mutant virus defective for nectin-1 binding through cells that express a gD-binding-impaired mutant nectin-1. The gB allele in a wild-type virus background enabled the use of other nectins as virus entry receptors. In addition, combination of the mutant allele with an epidermal growth factor receptor (EGFR)-retargeted gD gene yielded dramatically increased EGFR-specific virus entry compared to retargeted virus carrying wild-type gB. Entry of the gB mutant virus into nectin-1-bearing cells was markedly accelerated compared to that of wild-type virus, suggesting that the gB mutations affect a rate-limiting step in entry. Our observations indicate that ineffective gD activation can be complemented by hypersensitization of a downstream component of the entry cascade to gD signaling.
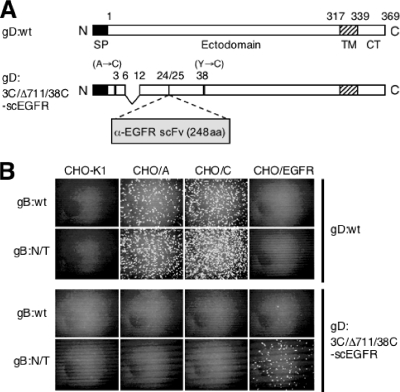
Figures
References
-
- Anderson, D. B., S. Laquerre, K. Ghosh, H. P. Ghosh, W. F. Goins, J. B. Cohen, and J. C. Glorioso. 2000. Pseudotyping of glycoprotein D-deficient herpes simplex virus type 1 with vesicular stomatitis virus glycoprotein G enables mutant virus attachment and entry. J. Virol. 74:2481-2487. - PMC - PubMed
-
- Asano, R., Y. Sone, K. Makabe, K. Tsumoto, H. Hayashi, Y. Katayose, M. Unno, T. Kudo, and I. Kumagai. 2006. Humanization of the bispecific epidermal growth factor receptor x CD3 diabody and its efficacy as a potential clinical reagent. Clin. Cancer Res. 12:4036-4042. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
